Мендель займався селекціонуванням гороху, і саме гороху, наукової удачі та суворості дослідів Менделя ми завдячуємо відкриттям основних законів успадкованості: закону однаковості гібридів першого покоління, закону розщеплення та закону незалежного комбінування.
Мендель експериментував з 22 різновидами гороху, що відрізнялися один від одного за 7 ознаками (колір, текстура насіння). Свою роботу Мендель вів вісім років, вивчив 10 тисяч рослин гороху. Усі форми гороху, що він досліджував, були представниками чистих ліній; результати схрещування таких рослин між собою завжди були однакові. Результати роботи Мендель навів у статті 1865, яка стала наріжним каменем генетики. Важко сказати, що заслуговує на більше захоплення в ньому і його роботі - строгість проведення експериментів, чіткість викладу результатів, досконале знання експериментального матеріалу або знання робіт його попередників.
Колеги та сучасники Менделя не змогли оцінити важливості зроблених ним висновків. За свідченням А.Є. Гайсиновича, до кінця ХІХ ст. її цитували лише п'ять разів, і лише один вчений – російський ботанік І.О. Шмальгаузен – оцінив важливість цієї роботи. Однак на початку XX століття закони, відкриті ним, були перевідкриті практично одночасно і незалежно один від одного вченими К. Корренсом, Е. Чермаком та К. де Фріз. Значимість цих відкриттів відразу стала очевидною науковому співтоваристві початку 1900-х років; їхнє визнання було пов'язане з певними успіхами цитології та формуванням гіпотези ядерної спадковості.
|
Основні закономірності успадкування ознак. Спадковість людини вивчена нині значно меншою мірою, ніж механізми успадкування ознак у рослинних і тваринних організмів. Проте вже сьогодні отримано цікаві дані про спадкову обумовленість багатьох фізичних ознак людини. Сучасний рівень генетичної науки дозволяє також стверджувати, що всі основні закономірності успадкування ознак та закони спадковості, виявлені в експериментах із рослинами та тваринами, є справедливими і для людини. Наприклад, якщо молодий чоловік, у якого в обох гомологічних хромосомах містяться гени, що забезпечують карий колір очей (гомозиготний за карим кольором), одружується з блакитноокою дівчиною, у якої в хромосомах також містяться тільки гени, які забезпечують блакитний колір очей, то їх потомство буде успадковувати колір очей за законами, відкритим засновником генетики Г. Менделем ще у 60-ті роки минулого століття у його дослідах із рослинними гібридами гороху. Спадкування статі в людини і більшості тварин також відбувається за загальним закономірностям і пов'язане з розподілом хромосом, що мають Х- і К-подібну форму (статеві хромосоми). У хромосомному наборі жіночого організму міститься дві Х-хромосоми, а чоловічому - одна хромосома X і одна У-хромосома (див. рис. 7). Число подібних прикладів може бути нескінченно великим, і немає сумніву, що «генний портрет» людини (генотип) визначає значною мірою багато його зовнішніх властивостей (фенотип). Нижче перераховані деякі спадкові ознаки людини, що визначаються домінантними та рецесивними генами (за К-Віллі, 1974). Прояв генного впливу може здійснюватися на різних етапах онтогенезу, але більшість фенотипічних ознак визначається ще до народження. Нарешті, і сам прояв генів немає фатальним, а залежить від чинників довкілля. Наприклад, важке спадкове захворювання фенілкетонурія не розвивається, якщо дитина, що містить генотипи гени, що викликають це захворювання, відразу ж після народження починає отримувати певну дієту. Таким чином, генетика має в своєму розпорядженні переконливі факти, що доводять існування як суворо обумовлених спадковістю ознак (наприклад, уппи крові, гемофілія, колір волосся і очей, риси обличчя та багато інших), так і ознак, що визначаються великою мірою зовнішнім середовищем (наприклад, зростання і маса людини, сила та спритність його м'язів, схильність до захворювань та ін.). Слід зазначити, що між генами та ознаками немає прямого зв'язку: розвиток однієї ознаки може залежати від впливу безлічі генів, а один ген може впливати на розвиток багатьох ознак. Ця картина ще більше ускладнюється постійною корекцією з боку довкілля. Отже, наїде венність _і з реай^каKjfcактотж^ розвитку завжди тісно взаємопов'язані. Не можна уявити розвиток дитини без коригуючих впливів середовища, як і неможливе без генетично зумовленої програми розвитку, що представляє синтез історичного досвіду всіх попередніх поколінь. Розглянемо роль спадковості та середовища у розвитку деяких фізичних ознак людини в процесі її пренатального та постнатального розвитку. Пренатальний розвиток. Формування органів прокуратури та функціональних систем дитини на процесі ембріогенезу перебуває під контролем генотипу, але чинники довкілля грають не останню роль. Для зародка первинним середовищем є материнський організм. Тут у спеціальному органі – матці – зародок відносно добре захищений від багатьох шкідливих впливів і отримує через плаценту все необхідне для його існування. Тим не менш, особливо на початкових етапах розвитку, багато факторів, що впливають на материнський організм, позначаються і на розвитку зародка (найбільш значними є: іонізуюче випромінювання, захворювання, перенесені жінкою під час вагітності, і багато хімічних речовин: алкоголь, нікотин, антибіотики, гормональні препарати та ін.). Слід зазначити, що в пренатальному періоді людини є критичні періоди, коли організм, що розвивається, особливо чутливий до дії зовнішніх факторів. Виділяють два такі періоди. Перший включає початок пренатального розвитку, його перші три тижні. У цей час відбувається закладання всіх найважливіших органів, і несприятливі дії у цей період частіше призводять до загибелі зародка. Протягом другого критичного періоду (з 4-го по 7-й тиждень) відбувається подальший розвиток усіх органів, і шкідливі зовнішні впливи в цей період можуть призвести до народження дитини з різними фізичними дефектами (рис. 9). Важливим є той факт, що однакові фізичні вроджені дефекти можуть бути пов'язані з пошкодженням генетичних структур, і з дією несприятливих зовнішніх факторів у процесі ембріогенезу. Це гарне свідчення того, що середовище та спадковість однаково відповідальні за нормальний розвиток дитини. Отже, на момент народження всі органи людини та її фізіологічні системи, зокрема і нервова система, виявляються загалом сформованими. Звичайно, виникає питання: чи є спадково зумовленими властивості нервової системи, які лежать в основі психічної діяльності людини, в основі її розуму? Чи є готові морфологічні «заготівлі свідомості» так само, як вони існують для нижчої нервової діяльності, пов'язаної з регуляцією життєвих процесів нашого тіла. Щоб відповісти на ці питання, зупинимося на наступному постнатальному етапі розвитку людини. Постнатальний розвиток дитини. Насамперед розглянемо основні методи, що дозволяють вивчати питоме значення середовища проживання і спадковості у постнатальном розвитку дітей та підлітків. Проведення подібних експериментів на рослинах є простим і полягає у виділенні двох груп організмів, ідентичних за генотипом, та приміщенні цих груп у різні умови існування. Наприклад, одну групу рослин (контрольну) вирощують у звичайних умовах, іншу (експериментальну) – у затемненому приміщенні. В результаті подібних дослідів можна зробити висновок, що утворення зеленого хлорофілу рослин залежить не тільки від спадковості, але і від факторів зовнішнього середовища (світла), оскільки рослини, що виросли в темряві, не міститимуть цей пігмент. Проведення аналогічних дослідів на людях неможливе як з морально-етичного боку, і з біологічної. У світі немає двох людей із однаковими генотипами. Але... є й винятки із правила. Це ідентичні, або гомозиготні, близнюки (ІБ), що мають не тільки вражаючу зовнішню схожість, а й майже однакові генні «портрети». Народження ідентичних та неідентичних близнюків (НБ) явище нерідке, в середньому зі 100 вагітностей одна завершується народженням більш ніж однієї дитини. Близнюки - чудовий «матеріал», подарований вченим самою природою, тому і сам метод називають близнюковим. У чому його суть? Припустимо, потрібно встановити, який фактор несе велику відповідальність за фізичний розвиток та зростання дитини: спадковість чи середовище? Для цього виділяють близнюкові пари, проводять їх морфологічне обстеження та вивчають спосіб життя (зазвичай близнюки, особливо ідентичні, мають подібні інтереси та близькі умови виховання). З проведених досліджень встановлюють коефіцієнт подібності (конкордантність) між близнюками і роблять висновок про роль спадковості чи середовища у розвитку тих чи інших якостей. Результати подібних досліджень ідентичних та неідентичних близнюків за частотою народження серед них деяких захворювань представлені в табл. На жаль, педагоги проводять поки що слабку роботу з виявлення обдарованих дітей, що часто пов'язано з недооцінкою дитячої фізіологічної та психічної індивідуальності, з незнанням елементарної біології дитини. Слабу роботу з виявлення дитячої обдарованості можна також пояснити ще низьким рівнем досліджень у цій галузі та відсутністю точних наукових критеріїв, за якими педагогові неважко було б виявити спадкові нахили дитини. Існує лише тісний зв'язок між обдарованістю та високим рівнем збудливості нервової системи, ознакою чого часто є різка неврівноваженість (психопатичність) дитини. До таких дітей педагог має ставитись особливо уважно. Це необхідно насамперед з гігієнічної сторони, оскільки неправильні дії педагога можуть призвести до розвитку крайніх характеристик характеру в патологічні. Необхідно це й у своєчасного виявлення спадкових задатків та його оптимального розвитку. Важливо пам'ятати, що видима відсутність схильностей до вчення, зовнішня льонність і недисциплінованість ще є достатніми доказами відсутності здібностей. З історії відомо чимало доказів справедливості цього висновку. Творець теорії відносності А. Ейнштейн у дитинстві не виявляв відкритих здібностей і лише у 9 років зміг піти у підготовчу школу. Погано навчалися в школі А. Гумбольдт (1769-1859) і Л. Пастер (1822-1895), видатний англійський фізик І. Ньютон мав славу в школі ледарем і нездатним учнем. Можна вважати, що вміле поводження з ними батьків та педагогів створило врешті-решт всі необхідні умови для розвитку талантів. Тим більше, що найважливішою якістю геніальності є працьовитість. Великий американський винахідник Т. Едісон писав: "Геній-це один відсоток натхнення, а на дев'яносто дев'ять відсотків потіння". Той, хто не навчиться «потіти», ніколи не зможе досягти великих успіхів у будь-якому вигляді людської діяльності, незважаючи на найвидатніші спадкові задатки. Хто, як не педагог, повинен насамперед виховувати у дитини працьовитість – цей найважливіший чинник прогресивного розвитку особистості, найголовніша умова формування будь-яких здібностей. Таким чином, спадковість лише дає «сирий матеріал», а середовище здійснює його основну «переробку». Людина, що народилася навіть із найсприятливішими задатками, але живе в середовищі, що перешкоджає розвитку її здібностей, залишиться посередністю, тож основна відповідальність за розвиток інтелектуальних можливостей дитини лягає на вихователів. Тим не менш роль середовища не слід абсолютизувати, не можна забувати, що «матеріал», що поставляється спадковістю, не завжди буває якісним і навіть найталановитіший скульптор не зможе з піску виліпити скульптуру, так само, як брила мармуру сама по собі не стане витвором мистецтва . №67 - №68. Аутосомно-домінантне успадкування- тип спадкування, при якому одного мутантного алелю, локалізованого в аутосомі, достатньо, щоб хвороба (або ознака) могла бути виражена. Аутосомно-рецесивне успадкування- тип успадкування ознаки або хвороби, при якому мутантний аллель, локалізований в аутосомі, має бути успадкований від обох батьків. |
№69. Спадкування, зчеплене із підлогою -успадкування будь-якого гена, що знаходиться в статевих хромосомах. Спадкування ознак, що виявляються тільки в особин одного статі, але не визначених генами, що знаходяться в статевих хромосомах, називається успадкуванням, обмеженою статтю.
Спадкуванням, зчепленим з X-хромосомою, називають успадкування генів у разі, коли чоловіча стать гетерогаметен і характеризується наявністю Y-хромосоми (XY), а особини жіночої статі гомогаметні і мають дві X-хромосоми (XX). Таким типом успадкування мають всі ссавці (у тому числі людина), більшість комах і плазунів.
№70 Голандричний тип успадкування.Голандричне успадкування (holandricinheritance) - успадкування ознаки, контрольованої геном, локалізованим у негомологічній частині Y-хромосоми. Голандричний ген (holandricgene) - ген, зчеплений із чоловічою статевою Y-хромосомою. Ознаки, що визначаються Р.р., передаються лише по чоловічій лінії – від батька до сина.
№71 При Х-зчепленому типі спадкуваннямутантний ген розташований у X-хромосомі. Якщо при цьому мутація має домінантний ефект, то хворими можуть бути як чоловіки, так і жінки. Однак від хворого батька захворювання з ймовірністю 100% передається лише дівчаткам, але не хлопчикам, які отримують від батька Y-хромосому (рис. 5). Імовірність передачі домінантної Х-зчепленої мутації від хворої матері дітям становить 50%. Хвороба з рівною ймовірністю може бути успадкована як дочкою, і сином.
До домінантного,зчепленому з Х-хромосомою, типу успадкування відноситься відома дитячим лікарям патологія вітамін Д-резистентний рахіт (синоніми: гіпофосфатемія, сімейна-Х-зчеплена гіпофосфатемія, фосфатдіабет). Діагноз цього найважчого рахіту, який не проходить під впливом великих доз вітаміну Д, підтверджується наявністю подібного захворювання у частини родичів як чоловічої, так і жіночої статі.
Набагато частіше Х-зчеплені захворюванняуспадковуються за рецесивним типом. Відмінною властивістю захворювань із рецесивнимХ-зчепленим типом успадкування і те, що у сім'ї хворіють чоловіки, а мутантний аллель вони успадковують від своєї, практично здорової матері, гетерозиготної по мутантному алелю. При складанні родоводу таких матерів нерідко спостерігаються хворі брати або дядьки. Хворі чоловіки можуть передавати своє захворювання лише через покоління і лише онукам (але не онукам) через свою здорову, але гетерозиготну дочку.
Таким чином, якщо простежити у родоводі успадкування по чоловічій лініїрецесивного Х-зчепленого захворювання, то вийде щось на кшталт «ходу шахового коня».
Найбільшвідомими Х-зчепленими захворюваннямиєгемофілії А та В , а також тяжка патологія м'язової системи -міодистрофіяДюшенна Беккера . В основі розвитку гемофілії А лежать мутації гена, відповідального за синтез VIII фактора згортання крові, а при гемофілії В дефектним виявляється IX фактор згортання крові. Обидва гени, що кодують фактори VIII і IX, локалізовані в довгому плечі Х-хромосоми в областях q28 та q27.1-2 відповідно. Відомо, що при гемофілії спостерігається порушення згортання крові, і найменші порізи можуть призвести хворого без спеціальної гематологічної допомоги до летального результату. Зазначимо, що у жінок - носіїв гена гемофілії (так званих «кондукторів»), в окремих випадках так само спостерігається схильність до кровотеч, що виражається у рясих місячних та тривалих кровотечах під час пологів. Цю обставину необхідно враховувати акушерам-гінекологам при роботі з жінками-носіями мутантних алелів у будь-якому гені гемофілії А або В.
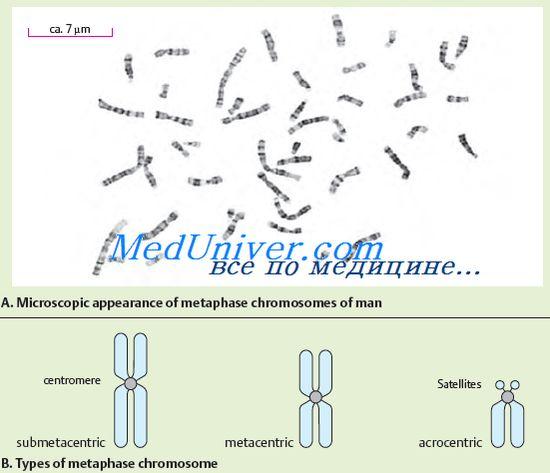
№72 Будова та функція хромосом. Морфологія хромосом найкраще видно у клітині на стадії метафази. Хромосома складається з двох паличкоподібних тілець – хроматид. Обидві хроматиди кожної хромосоми ідентичні один одному генним складом. Хромосоми диференційовані за довжиною. Хромосоми мають центромір або первинну перетяжку, дві теломери і два плечі. На деяких хромосомах виділяють вторинні перетяжки та супутники. Рух хромосоми визначає Центромера, що має складну будову. ДНК центроміри відрізняється характерною послідовністю нуклеотидів та специфічними білками. Залежно від розташування центроміри розрізняють акроцентричні, субметацентричні та метацентричні хромосоми. Як говорилося вище, деякі хромосоми мають вторинні перетяжки. Вони, на відміну від первинної перетяжки (центроміри), не є місцем прикріплення ниток веретена і не відіграють жодної ролі в русі хромосом. Деякі вторинні перетяжки пов'язані із заснуванням ядерців, у разі їх називають ядерцевими організаторами. У ядерцевих організаторах розташовані гени, відповідальні за синтез РНК. Функція інших вторинних перетяжок ще зрозуміла. У деяких акроцентричних хромосом є супутники - ділянки, з'єднані з рештою хромосоми тонкою ниткою хроматину. Форма та розміри супутника постійні для цієї хромосоми. У людини супутники є п'ять пар хромосом. Кінцеві ділянки хромосом, багаті на структурний гетерохроматин, називаються теломерами. Теломери перешкоджають злипання кінців хромосом після редуплікації і тим самим сприяють збереженню їхньої цілісності. Отже, теломери відповідальні існування хромосом як індивідуальних утворень. Хромосоми, що мають однаковий порядок генів, називають гомологічними. Вони мають однакову будову (довжина, розташування центроміри тощо). Негомологічні хромосоми мають різний генний набір та різну будову. Дослідження тонкої структури хромосом показало, що вони складаються з ДНК, білка та невеликої кількості РНК. Молекула ДНК несе негативні заряди, розподілені по всій довжині, а приєднані до неї білки – гістони заряджені позитивно. Цей комплекс ДНК із білком називають хроматином. Хроматин може мати різний ступінь конденсації. Конденсований хроматин називають гетерохроматином, деконденсований хроматин – еухроматином. Ступінь деконденсації хроматину відбиває його функціональний стан. Гетерохроматинові ділянки функціонально менш активні, ніж еухроматинові, у яких локалізована більшість генів. Розрізняють структурний гетерохроматин, кількість, якого різниться у різних хромосомах, але розташовується він у навколоцентромірних районах. Крім структурного гетерохроматину, існує факультативний гетерохроматин, який з'являється в хромосомі при надспіралізації еухроматичних районів. Підтвердженням існування цього явища в хромосомах людини є факт генетичної інактивації однієї Х-хромосоми в соматичних клітинах жінки. Його суть полягає в тому, що існує механізм інактивації другої дози генів, що еволюційно сформувався, генів, локалізованих в Х-хромосомі, внаслідок чого, незважаючи на різне число Х-хромосом в чоловічому і жіночому організмах, число функціонуючих в них генів зрівняно. Максимально конденсований хроматин під час мітотичного поділу клітин, тоді його можна виявити у вигляді щільних хромосом. Розміри молекул ДНК хромосом величезні. Кожна хромосома представлена однією молекулою ДНК. Вони можуть сягати сотень мікрометрів і навіть сантиметрів. З хромосом людини найбільша – перша; її ДНК має загальну довжину до 7 см. Сумарна довжина молекул ДНК усіх хромосом однієї клітини людини становить 170 см. Незважаючи на гігантські розміри молекул ДНК, вона досить щільно запакована в хромосомах. Таке специфічне укладання хромосомної ДНК забезпечують білки гістони. Гістони розташовуються за довжиною молекули ДНК як блоків. В один блок входить 8 молекул гістонів, утворюючи нуклеосому (освіта, що складається з нитки ДНК намотаної навколо октамера гістонів). Розмір нуклеосоми близько 10 нм. Нуклеосоми мають вигляд нанизаних на нитку бусинок. Нуклеосоми і ділянки ДНК, що їх з'єднують, щільно упаковані у вигляді спіралі, на кожен виток такої спіралі припадає шість нуклеосом. Так формується структура хромосоми. Спадкова інформація організму суворо впорядкована за окремими хромосомами. Кожен організм характеризується певним набором хромосом (число, розміри та структура), який називається каріотипом. Каріотип людини представлений двадцятьма чотирма різними хромосомами (22 пари аутосом, Х- та Y-хромосоми). Каріотип – це паспорт виду. Аналіз каріотипу дозволяє виявляти порушення, які можуть призводити до аномалій розвитку, спадкових хвороб або загибелі плодів та ембріонів на ранніх стадіях розвитку. Довгий час вважали, що каріотип людини складається з 48 хромосом. Однак на початку 1956 було опубліковано повідомлення, згідно з яким число хромосом в каріотипі людини дорівнює 46. Хромосоми людини різняться за розміром, розташуванням центроміри і вторинних перетяжок. Вперше підрозділ каріотипу на групи було проведено 1960 р. на конференції в Денвері (США). В опис каріотипу людини спочатку було закладено два наступні принципи: розташування хромосом за їх довжиною; угруповання хромосом за розташуванням центроміри (метацентричні, субметацентричні, акроцентричні). Точна сталість числа хромосом, їх індивідуальність і складність будови свідчать про важливість виконуваної ними функції. Хромосоми виконують функцію головного генетичного апарату клітини. Вони в лінійному порядку розташовані гени, кожен із яких займає строго певне місце (локус) в хромосомі. У кожній хромосомі багато генів, але нормального розвитку організму необхідний набір генів повного хромосомного набору.
№73. В основі спадковості лежить здатність всіх живих організмів накопичувати, зберігати та передавати потомству спадкову інформацію. Ця одна з найважливіших якісних особливостей живої матерії пов'язана з нуклеїновими кислотами - дезоксирибонуклеїнової (ДНК) та рибонуклеїнової (РНК). Провідне значення належить ДНК - найдовшій молекулі живих організмів, зосередженої в ядрах клітин і її спадковий апарат. Велика довжина молекули ДНК дає можливість "записати" на ній, як на телеграфній стрічці, всі основні властивості майбутнього організму та програму його розвитку. Такий «запис» здійснюється за допомогою спеціальної «нуклеїнової мови», або «нуклеїнового коду», сутністю якого є зміна порядку проходження чотирьох хімічних сполук, що входять до складу ДНК. та цілі пропозиції «нуклеїнової мови».
На такій нуклеїновій «стрічці» можна виділити окремі самостійні ділянки, що включають опис програми розвитку однієї ознаки. Їх називають генами. Кожна молекула ДНК включає сотні генів і є програмою розвитку багатьох ознак і властивостей організму. Поєднуючись з особливими білками та деякими іншими речовинами, молекули ДНК утворюють у ядрі спеціальні освіти - звані хромосоми.
Організація генетичного матеріалу. Гени [від грец. genos, народження] - одиниця спадковості, ділянка ДНК, що займає специфічне місце у хромосомі. З погляду генетики, ген - успадкований чинник і неподільна одиниця генетичного матеріалу. Структурний ген (цистрон) – фрагмент ДНК, що бере участь у освіті поліпептидного ланцюга. До його складу входять лідерна послідовність, що кодують фрагменти (екзони), вставні послідовності (нітрони) та кінцева послідовність. Оскільки деякі білки складаються з більш ніж однієї субодиниці, формулювання «один ген - один фермент» стосовно гетеромультимерного (тобто що складається з двох і більше різних поліпептидних субодиниць) білку слід трактувати як «один ген - один поліпептидний ланцюг».
Генотип – сукупність генів організму. Ще в давнину люди емпірично використовували закономірності спадкування. На підставі цього досвіду набула розвитку селекція [від лат. selectio, обирати] - наука про методи створення нових сортів рослин та порід тварин шляхом відбору та схрещування. Донедавна генотип здавався неприступним, не підвладним діям людини. Відкриття структури генів дозволило виділяти в ізольованому вигляді, синтезувати біохімічно і навіть вводити в організм. Стало можливим вплив на ген без виділення з організму. Усе це створило передумови маніпулювання генотипом.
№74 Будова нуклеїнових кислот.
Нуклеїнова кислота(від лат. Nucleus - ядро) - високомолекулярна органічна сполука, біополімер (полінуклеотид), утворений залишками нуклеотидів. Нуклеїнові кислотиДНКіРНК присутні у клітинах всіх живих організмів і виконують найважливіші функції зі зберігання, передачі та реалізації спадкової інформації. Полімерні форми нуклеїнових кислот називають полінуклеотидами. Ланцюжки з нуклеотидів з'єднуються через залишок фосфорної кислоти (фосфодіефірний зв'язок). Оскільки в нуклеотидах існує тільки два типи гетероциклічних молекул, рибоза і дезоксирибозу, то є лише два види нуклеїнових кислот -дезоксирибонуклеїнова (ДНК) ірибонуклеїнова (РНК).
Мономірні форми також зустрічаються в клітинах та відіграють важливу роль у процесах передачі сигналів або запасанні енергії. Найбільш відомий мономер РНК – АТФ, аденозинтрифосфорна кислота, найважливіший акумулятор енергії у клітині.
Одна з основних функцій нуклеїнових кислот полягає у детермінації синтезу білків. Інформація про структуру білків, закодована в нуклеотидній послідовності ДНК, повинна передаватися від одного покоління до іншого, і тому необхідне безпомилкове копіювання, тобто. синтез такої самої молекули ДНК (реплікація).
№75Ріплікація ДНК- процес синтезу дочірньої молекули дезоксирибонуклеїнової кислоти на матриці батьківської молекули ДНК. У ході подальшого поділу материнської клітини кожна дочірня клітина отримує по одній копії молекули ДНК, яка є ідентичною ДНК вихідної материнської клітини. Цей процес забезпечує точну передачу генетичної інформації з покоління до покоління. Реплікацію ДНК здійснює складний ферментний комплекс, що складається з 15-20 різних білків, званий реплісомою (англ. replisome). Реплікація ДНК - ключова подія в ході поділу клітини. Принципово, щоб на момент поділу ДНК була реплікована повністю і лише один раз. Це забезпечується певними механізмами регулювання реплікації ДНК. Реплікація відбувається у три етапи:
ініціація реплікації
елонгація
Термінація реплікації.
Регуляція реплікації здійснюється переважно на етапі ініціації. Це досить легко здійснимо, тому що реплікація може починатися не з будь-якої ділянки ДНК, а з певного, званого сайтом ініціації реплікації. У генометаких сайтів може бути як всього один, так і багато. З поняттям сайту ініціації реплікації тісно пов'язане поняття реплікон. Реплікон - це ділянка ДНК, яка містить сайт ініціації реплікації та реплікується після початку синтезу ДНК із цього сайту. Геномибактерій, як правило, є одним репліконом, це означає, що реплікація всього геному є наслідком всього одного акта ініціації реплікації. Геномиеукаріот (а також їх окремі хромосоми) складаються з великої кількості самостійних репліконів, це значно скорочує сумарний час реплікації окремої хромосоми. Молекулярні механізми, які контролюють кількість актів ініціації реплікації в кожному сайті за один цикл розподілу клітини, називаються контролем копійності. У бактеріальних клітинах крім хромосомної ДНК часто містяться плазміди, які являють собою окремі реплікони. У плазмід існують свої механізми контролю копійності: вони можуть забезпечувати синтез як однієї копії плазміди заклітинний цикл, так і тисяч копій.
Реплікація починається в сайті ініціації реплікації з розплетення подвійної спіралі ДНК, при цьому формується реплікаційна вилка - місце безпосередньої реплікації ДНК. У кожному сайті може формуватися одна чи дві реплікаційні вилки залежно від того, чи є реплікація одно- чи двонаправленою. Найбільш поширена двонаправлена реплікація. Через деякий час після початку реплікації в електронний мікроскоп можна спостерігати реплікаційне вічко - ділянку хромосоми, де ДНК вже реплікована, оточений більш протяжними ділянками нереплікованої ДНК.
У реплікаційній вилці ДНК копіює великий білковий комплекс (реплісома), ключовим ферментом якого є ДНК-полімераза. Реплікаційна вилка рухається зі швидкістю близько 100 000 пар нуклеотидів за хвилину у прокаріотів і 500-5000 - у еукаріотів.
№76 Будова та види хромосом.У ядрі кожної соматичної клітини організму людини міститься46 хромосом. Набірхромосом кожного індивідуума, як нормальний, і патологічний, називається каріотипом.
З46 хромосом, складових хромосомний набір людини, 44 або 22 пари представляють аутосомніхромосоми , остання пара - статеві хромосоми У жінок конституція статевих хромосом у нормі представлена двома хромосомами X, а у чоловіків – хромосомами X та У.
У всіхпарах хромосомяк аутосомних, так і статевих одна з хромосом отримана від батька, а друга – від матері. Хромосоми однієї пари називаються гомологами або гомологічними хромосомами. У статевих клітинах (сперматозоїдах та яйцеклітинах) міститься гаплоїдний набір хромосом, тобто. 23 хромосоми.
 Сперматозоїди поділяються на два типи залежно від того, чи містять вонихромосомуX або Y. Усі яйцеклітини в нормі містятьтільки хромосому
X.
Сперматозоїди поділяються на два типи залежно від того, чи містять вонихромосомуX або Y. Усі яйцеклітини в нормі містятьтільки хромосому
X.
Хромосомидобре видно після спеціального забарвлення під час поділу клітин, коли хромосоми максимально спіралізовані. При цьому в кожній хромосомі виявляється перетяжка, яка називається центроміром. Центромера ділить хромосому на коротке плече (позначається літерою "р") та довге плече (позначається літерою "q"). Центромера визначає рух хромосоми під час клітинного поділу. За положенням центроміри хромосоми класифікують на кілька груп. Якщо центроміра розташовується посередині хромосоми, то така хромосома називається метацентричної, якщо центроміра розташовується ближче до одного з кінців хромосоми, то її називають акроцентричної. Деякі акроцентричні хромосоми мають так звані супутники, які в клітині, що не ділиться, формують ядерця. Ядерця містять численні копії рРНК. Крім того, розрізняють субметацентричні хромосоми, коли центроміра розташована не посередині хромосоми, а дещо зсунута до одного з кінців, але не настільки значно, як в акроцентричних хромосомах.
Кінці кожногоплеча хромосоминазивають теломірами. Встановлено, що теломери відіграють важливу роль у збереженні хромосом стабільності. У теломерах міститься велика кількість повторів послідовності нуклеотидів ТТАГГГ, про тандемних повторів. У нормі під час клітинного поділу відбувається зменшення числа цих повторів у тіломірах .
Однак щоразу вони добудовуються за допомогою спеціального ферменту, який називаютьтіломіразою. Зменшення активності цього ферменту призводить до скорочення теломер, що, як вважають, є причиною загибелі клітин, а в нормі супроводжує старіння.
№77. Денверська класифікація хромосом.Класифікація і номенклатура рівномірно забарвлених хромосом людини вперше були прийняті на міжнародній нараді в 1960 році в Денвері, надалі дещо змінені і доповнені (Лондон, 1963 і Чикаго, 1966). Згідно з Денверівською класифікацією всі хромосоми людини розділені на 7 груп, розташованих у порядку зменшення їх довжини та з урахуванням центріольного індексу (відношення довжини короткого плеча до довжини всієї хромосоми, виражене у відсотках). Групи позначаються буквами англійського алфавіту від А до G. Усі пари хромосом прийнято нумерувати арабськими цифрами. Характеристика груп представлена у табл. 4. Запропонована класифікація дозволяла чітко розрізняти хромосоми, що належать до різних груп. З 1960 починається бурхливий розвиток клінічної цитогенетики: в 1959 Дж. Лежен відкрив хромосомну природу синдрому Дауна; К. Форд, П. Джекобс та Дж. Стронг описали особливості каріотипу при синдромах Клайнфельтера та Тернера; на початку 70-х років. було відкрито хромосомну природу синдромів Едвардса, Патау, синдрому «котячого крику»; описана хромосомна нестабільність при низці спадкових синдромів та злоякісних захворюваннях. Водночас застосування методу одержання рівномірно забарвлених хромосом виявилося недостатньо ефективним для ідентифікації хромосом. Недоліком денверської класифікації і те, що розмежування гомологічних пар усередині групи хромосом зустрічає найчастіше непереборні труднощі.
Таблиця 4^ Групи хромосом та їх характеристика
|
Група |
№ хромосоми |
Розташування центроміру |
Центріольний індекс (%) |
Примітка |
|
Найбільша метацентрична |
На довгому плечі може бути вторинна перетяжка |
|||
|
Найбільша субметацентрична | ||||
|
Велика метацентрична |
На 20% коротше за першу |
|||
|
Велика субметацентрична | ||||
|
6-12 та Х-хромосома |
Середні субметацентричні |
На 9-й часто вторинна перетяжка |
||
|
Середні акроцентричні |
На всіх вторинні перетяжки |
|||
|
Маленька метацентрична |
У 10% випадків зустрічається вторинна перетяжка |
|||
|
Маленька субметацентрична | ||||
|
Найменші метацентричні | ||||
|
21-22 та Y-хромосома |
Найменші акроцентричні |
На 21-й та 22-й вторинні перетяжки |
№78 Біосинтез білка у клітині.Процес синтезу білка є складним багатоступеневим процесом. Здійснюється він у спеціальних органелах – рибосомах. У клітині міститься велика кількість рибосом. Наприклад, у кишкової палички їх близько 20 тисяч.
Як відбувається синтез білка в рибосомах?
Молекули білків по суті є поліпептидні ланцюжки, складені з окремих амінокислот. Але амінокислоти недостатньо активні, щоб поєднатися між собою самостійно. Тому, перш ніж з'єднатися один з одним та утворити молекулу білка, амінокислоти повинні активуватися. Ця активація відбувається під впливом спеціальних ферментів. Причому кожна амінокислота має свій, специфічно налаштований її у фермент.
Джерелом енергії для цього (як і для багатьох процесів у клітині) є аденозинтрифосфат (АТФ).
Внаслідок активування амінокислота стає більш лабільною і під дією того ж ферменту зв'язується з т-РНК.
Важливим є те, що кожній амінокислоті відповідає суворо специфічна т-РНК. Вона знаходить свою амінокислоту і переносить її в рибосому. Тому така РНК отримала назву транспортної.
Отже, рибосому надходять різні активовані амінокислоти, з'єднані зі своїми т-РНК. Рибосома являє собою ніби конвеєр для складання ланцюжка білка з різних амінокислот, що надходять до нього.
Виявляється, що одночасно з т-РНК, на якій «сидить» своя амінокислота, рибосому надходить «сигнал» від ДНК, яка міститься в ядрі. Відповідно до цього сигналу в рибосомі синтезується той чи інший білок, той чи інший фермент (оскільки ферменти є білками).
Напрямний вплив ДНК на синтез білка здійснюється не безпосередньо, а за допомогою особливого посередника тієї форми РНК, яка отримала назву матричної або інформаційної РНК (м-РНК або і-РНК).
Інформаційна РНК синтезується в ядрі йод впливом ДНК, тому її склад відбиває склад ДНК. Молекула РНК є як би зліпок з форми ДНК.
Синтезована і-РНК надходить у рибосому і як би передає цій структурі план - в якому порядку повинні з'єднуватися один з одним активовані амінокислоти, що надійшли в рибосому, щоб синтезувався певний білок. Інакше генетична інформація, закодована в ДНК, передається на і-РНК і далі на білок.
№79 Мутація (лат.mutatio - зміна) -стійка (тобто така, яка може бути успадкована нащадками даної клітини або організму) зміна генотипу, що відбувається під впливом зовнішнього або внутрішнього середовища. Термін запропонований Гуго де Фрізом. Процес виникнення мутацій отримав назву мутагенезу.
Геномні: - поліплоїдизація (освіта організмів або клітин, геном яких представлений більш ніж двома (3n, 4n, 6n і т. д.) наборами хромосом) і анеуплоїдія (гетероплоїдія) - зміна числа хромосом, не кратна гаплоїдному набору (див. Інге-Вечтомов , 1989). Залежно від походження хромосомних наборів серед поліплоїдів розрізняють алолополіплоїди, у яких є набори хромосом, отримані при гібридизації від різних видів, і аутополіплоїдів, у яких відбувається збільшення числа наборів хромосом власного геному, кратне n.
При хромосомних мутаціях відбуваються великі перебудови структури окремих хромосом. У цьому випадку спостерігаються втрата (делеція) або подвоєння частини (дуплікація) генетичного матеріалу однієї або декількох хромосом, зміна орієнтації сегментів хромосом в окремих хромосомах (інверсія), а також перенесення частини генетичного матеріалу з однієї хромосоми на іншу (транслокація) (крайній випадок - об'єднання цілих хромосом, т. зв. Робертсонівська транслокація, яка є перехідним варіантом від хромосомної мутації до геномної).
На генному рівні зміни первинної структури ДНК генів під дією мутацій менш значні, ніж при хромосомних мутаціях, проте генні мутації зустрічаються частіше. В результаті генних мутацій відбуваються заміни, делеції та вставки одного або декількох нуклеотидів, транслокації, дуплікації та інверсії різних частин гена. У тому випадку, коли під дією мутації змінюється лише один нуклеотид, говорять про точкові мутації. Оскільки до складу ДНК входять азотисті підстави лише двох типів - пурини і піримідини, всі точкові мутації із заміною основ поділяють на два класи: транзиції (заміна пурину на пурин або піримідину на піримідин) і трансверсії (заміна пурину на піримідин або навпаки). Можливі чотири генетичні наслідки точкових мутацій: 1) збереження сенсу кодона через виродженість генетичного коду (синонімічна заміна нуклеотиду); 2) зміна сенсу кодону, що призводить до заміни амінокислоти у відповідному місці поліпептидного ланцюга термінацією (нонсенс-мутація). У генетичному коді є три безглузді кодони: амбер - UAG, охр - UAA і опал - UGA (відповідно до цього отримують назву і мутації, що призводять до утворення безглуздих триплетів - наприклад амбер-мутація), 4) зворотна заміна (стоп-кодона на смисловий кодон).
За впливом на експресію генів мутації поділяють на дві категорії: мутації типу замін пар основ та типу зсуву рамки зчитування (frameshift). Останні є делеції або вставки нуклеотидів, число яких не кратно трьом, що пов'язано з триплетністю генетичного коду.
Первинну мутацію іноді називають прямою мутацією, а мутацію, що відновлює вихідну структуру гена, - зворотною мутацією, або реверсією. Повернення до вихідного фенотипу мутантного організму внаслідок відновлення функції мутантного гена нерідко відбувається не за рахунок істинної реверсії, а внаслідок мутації в іншій частині того самого гена або навіть іншого неалельного гена. І тут зворотну мутацію називають супрессорной. Генетичні механізми, завдяки яким відбувається супресія мутантного фенотипу, дуже різноманітні.
Ниркові мутації (спори) - стійкі соматичні мутації, що відбуваються в клітинах точок росту рослин. Приводять до клонової мінливості. При вегетативному розмноженні зберігаються. Багато сортів культурних рослин є нирковими мутаціями.
№80. Упаковка ДНК у хромосому.Хромосоми складаються з хроматину - суміші ДНК та білків у співвідношенні ~ 1:1. Саме завдяки взаємодії із білками молекула ДНК має компактну структуру. Середня довжина однієї нитки ДНК людини (якщо розтягнути) 4 див. Тобто ядрі кожної клітини міститься близько 1 метра ДНК. Упаковка молекули ДНК у хромосомі має три рівні (див. малюнок): 1.нуклеосоми, нуклеосомна нитка 2. соленоїдна спіраль 3. петлі Відома кожному своїм виглядом подвійна спіраль ДНК «намотанна» навколо комплексу з 4-х пар білкових молекул утворює «бусину» - Нуклеосому. Нуклеосоми, пов'язані між собою ділянками молекули ДНК, складають нитку, яка у свою чергу закручена у формі соленоїда (це за однією з існуючих моделей), один виток – 6 нуклеосом. Ця структура наступного рівня упаковки утворює петлі. У такому вигляді ДНК є в ядрі клітини на етапі росту клітини, синтезу її компонентів, у т. ч. подвоєння ДНК. Однак, на різних стадіях життя клітини конденсація хроматину різна. У період зростання клітини хроматин у ядрі не конденсовано, компактність його розташування при цьому відрізняється для різних ділянок хромосоми. Коли відбувається процес поділу клітини хроматин дедалі більше ущільнюється і петлі, про які йшлося вище, утворюють «розетки», щільно укладені одна до одної (теж по одній з існуючих моделей), по 18 петель на кільце. В результаті цього хроматин набуває вигляду конденсованих хромосом, так добре відомий нам з підручників.
№81. Гаметогенез.(від Гамети та грец. genesis - походження) процес розвитку та формування статевих клітин - гамет. Г. чоловічих гамет (сперматозоїдів, сперміїв) називають сперматогенезом, жіночих гамет (яйцеклітин) -оогенезом. У тварин і рослин Р. протікає по-різному, залежно від місця Мейоза в життєвому циклі цих організмів.
У багатоклітинних тварин Р. відбувається в спеціальних органах - статевих залозах, або гонадах (яєчниках, сім'яниках, гермафродитних статевих залозах), і складається з трьох основних етапів: 1) розмноження первинних статевих клітин - гаметогоніїв (сперматогоніїв і оогоніїв) шляхом ряду 2) зростання і дозрівання цих клітин, називають тепер гаметоцитами (сперматоцитами і ооцитами), які, як і гаметогонії, мають повний (здебільшого диплоїдний) набор хромосом (Див. Хромосоми). У цей час відбувається основна подія Р. у тварин - розподіл гаметоцитів шляхом мейозу, що призводить до редукції (зменшення вдвічі) числа хромосом у цих клітинах і перетворення їх на гаплоїдні клітини (см. Гаплоїд) - сперматиди та оотиди; 3) формування сперматозоїдів (або сперміїв) та яйцеклітин; при цьому яйцеклітини одягаються поруч зародкових оболонок, а сперматозоїди набувають джгутики, що забезпечують їх рухливість. У самок багатьох видів тварин мейоз та формування яйця завершуються після проникнення сперматозоїда в цитоплазму ооциту, але до злиття ядер сперматозоїда та яйцеклітини.
У рослин Р. відокремлений від мейозу і починається в гаплоїдних клітинах – у суперечках (у вищих рослин – мікроспори та мегаспори). Зі суперечок розвивається статеве покоління рослини - гаплоїдний Гаметофіт, у статевих органах якого - гаметангії, жіночих - архегоніях шляхом мітозів відбувається Г. Виняток становлять голонасінні і покритонасінні рослини, у яких статеві рослини йде безпосередньо в проростаючій мікроспорі - пилковій клітині. У всіх нижчих і вищих спорових рослин Р. в антеридиях - це багаторазове розподіл клітин, у результаті якого утворюється велика кількість дрібних рухомих сперматозоїдів. Р. в архегоніях - формування однієї, двох або кількох яйцеклітин. У голонасінних і покритонасінних рослин чоловічий Р. складається з поділу (шляхом мітозу) ядра пилкової клітини на генеративний і вегетативний і подальшого поділу (також шляхом мітозу) генеративного ядра на два спермію. Цей поділ відбувається в пилковій трубці, що проростає. Жіночий Р. у покритонасінних рослин - відокремлення шляхом мітозу однієї яйцеклітини всередині 8-ядерного зародкового мішка. Основна відмінність Р. у тварин і рослин: у тварин він поєднує в собі перетворення клітин з диплоїдних на гаплоїдні та формування гаплоїдних гамет; у рослин Р. зводиться до формування гамет із гаплоїдних клітин.
№82 Каріотипсукупність ознак (число, розміри, форма і т. д.) повного набору хромосом, властива клітинам даного біологічного виду (видовий каріотип), даного організму (індивідуальний каріотип) або лінії (клону) клітин. Каріотипом іноді називають і візуальне подання повного хромосомного набору (каріограми). Зовнішній вигляд хромосом істотно змінюється протягом клітинного циклу: протягом інтерфази хромосоми локалізовані відер, як правило, деспіралізовані і важкодоступні для спостереження, тому для визначення каріотипу використовуються клітини в одній зі стадій їх поділу -метафазі мітозу. Для процедури визначення каріотипу можуть бути використані будь-які популяції клітин, що діляться. Для визначення людського каріотипу використовують, як правило, лімфоцити периферичної крові, перехід яких від стадії спокою G0 до проліферації провокують додаванням мітогенафітогемагглютініна. Для визначення каріотипу можуть бути використані також клітини кісткового мозку або первинна культура фібробластів шкіри. Для збільшення числа клітин на стадії метафазик культурі клітин незадовго перед фіксацією додають колхіцинілінокадазол, які блокують утворення мікротрубочок, тим самим перешкоджаючи розбіжності хроматид до полюсів поділу клітини і завершення мітозу.
Після фіксації препарати метафазних хромосом забарвлюють та фотографують; з мікрофотографій формують так званий систематизований каріотип - нумерований набір пар гомологічних хромосом, зображення хромосом при цьому орієнтуються вертикально короткими плечима вгору, їх нумерація проводиться в порядку зменшення розмірів, пара статевих хромосом поміщається в кінець набору (див. Рис .
Історично перші недеталізовані каріотипи, що дозволяли проводити класифікацію з морфології хромосом, отримували забарвленням по Романовському - Гімзі, проте подальша деталізація структури хромосом у каріотипах стала можливою з появою методик диференціального фарбування хромосом. Найбільш часто використовуваною методикою в медичній генетиці є метод G-диференціального фарбування хромосом.
Порівняння комплексів поперечних міток у класичній каріотипії або ділянок зі специфічними спектральними характеристиками дозволяє ідентифікувати як гомологічні хромосоми, так і окремі їх ділянки, що дозволяє детально визначати хромосомні аберації-внутрішньо- та міжхромосомні перебудови, що супроводжуються порушенням порядку фрагментів, транслокації). Такий аналіз має велике значення в медичній практиці, дозволяючи діагностувати ряд хромосомних захворювань, спричинених як грубими порушеннями каріотипів (порушення числа хромосом), так і порушенням хромосомної структури або множинністю каріотипів клітин в організмі (мозаїцизмом).
№83Мейоз- редукційний поділ, поділ дозрівання, спосіб поділу клітин, в результаті якого відбувається зменшення (редукція) числа хромосом в два рази і одна диплоїдна клітина (що містить два набори хромосом) після двох швидко ділянок, що швидко наступають один за одним, дає початок 4 гаплоїдним (що містить по одному набору хромосом).
Біологічне значення М . полягає у підтримці сталостіКаріотипа в ряді поколінь організмів даного виду та забезпечення можливості рекомбінації) хромосом і генів при статевому процесі. М. – один із ключових механізмів спадковості
Мейоз складається з 2 послідовних поділів із короткою інтерфазою між ними.
Профаза I – профаза першого поділу дуже складна і складається з 5 стадій:
Лептотена чи лептонема - упаковка хромосом, конденсація ДНК із заснуванням хромосом як тонких ниток (хромосоми коротшають).
Зиготена або зигонема - відбувається кон'югація - з'єднання гомологічних хромосом з утворенням структур, що складаються з двох сполучених хромосом, званих зошитами або бівалентами та їх подальша компактизація.
Пахітена або пахінема – (найдовша стадія) – у деяких місцях гомологічні хромосоми щільно з'єднуються, утворюючи хіазми. У них відбувається кросинговер- обмін ділянками між гомологічними хромосомами.
Диплотена чи диплонема - відбувається часткова деконденсація хромосом, у своїй частина геному може працювати, відбуваються процеси транскрипції (освіта РНК), трансляції (синтез білка); гомологічні хромосоми залишаються з'єднаними між собою. У деяких тварин в ооцитах хромосоми на цій стадії профази мейозу набувають характерної форми хромосом типу лампових щіток.
Діакінез – ДНК знову максимально конденсується, синтетичні процеси припиняються, розчиняється ядерна оболонка; центріолі розходяться до полюсів; гомологічні хромосоми залишаються з'єднаними між собою.
До кінця Профази I центріолі мігрують до полюсів клітини, формуються нитки веретена поділу, руйнуються ядерна мембрана та ядерця
Метафаза I - бівалентні хромосоми вишиковуються вздовж екватора клітини.
Анафаза I – мікротрубочки скорочуються, біваленти діляться і хромосоми розходяться до полюсів. Важливо відзначити, що через кон'югацію хромосом у зиготені до полюсів розходяться цілі хромосоми, що складаються з двох хроматид кожна, а не окремі хроматиди, як у мітоз.
Телофаза I – хромосоми деспіралізуються та з'являється ядерна оболонка.
Другий поділ мейозу слідує безпосередньо за першим, без вираженої інтерфази: S-період відсутній, оскільки перед другим розподілом не відбувається реплікації ДНК.
Профаза II - відбувається конденсація хромосом, клітинний центр ділиться і продукти його поділу розходяться до полюсів ядра, руйнується ядерна оболонка, утворюється веретено поділу.
Метафаза II - унівалентні хромосоми (що складаються з двох хроматид кожна) розташовуються на «екваторі» (на рівній відстані від «полюсів» ядра) в одній площині, утворюючи так звану метафазну пластинку.
Анафаза II - уніваленти діляться іхроматидирозходяться до полюсів.
Телофаза II – хромосоми деспіралізуються та з'являється ядерна оболонка.
В результаті з однієїдиплоїдної клітиниутворюється чотиригаплоїдні клітини. У тих випадках, коли мейоз пов'язаний згаметогенезом(наприклад, у багатоклітинних тварин), при розвиткуяйцеклітинперше і друге поділу мейозу різко нерівномірні. У результаті формується одна гаплоїдна яйцеклітина та три так званих редукційних тільця(Абортивні деривати першого та другого поділів).
№84 Розподіл клітин – мітоз
Якби ви спостерігали під мікроскопом за розподілом клітин, то побачите наступне. По-перше, матеріал усередині ядра стає дуже щільним. Можна навіть розрізнити дискретні структури – хромосоми. Ви знаєте, що в соматичних (нестатевих) клітинах людини є 23 пари хромосом – по одній хромосомі з кожної пари від кожного з батьків. Разом у соматичних клітинах людини 46 хромосом. Усі 46 вишиковуються в екваторіальній площині клітини. На цій стадії добре помітно, що кожна з 46 хромосом насправді складається з двох хроматид, з'єднаних між собою області, схожій на кнопку. Цю область було названо центроміром. З протилежних кінців клітини розташовано дві щільні плями (центріолі), кожна з яких утворює нитки веретена поділу. Кожна нитка приєднується до хромосоми в області центроміру. До кожної пари хромосом приєднуються дві нитки – по одній від кожної центріолі. Потім ми б побачили, що центріолі, подібно до котушок, намотують нитки веретена поділу, внаслідок чого хромосоми розходяться до полюсів клітини. Потім ділиться вміст клітини. Це дивовижне видовище.
Події розвиваються за наступним сценарієм. Необхідно розуміти, що для поділу клітини має бути вирішено дуже важливе завдання - має відбутися подвоєння (реплікація) ДНК, адже саме в ДНК закодовано всю інформацію про те, що і як має відбуватися в клітині. Згадайте, що ДНК є подвійною спіралью. Зовні спіралі розташовується сахарофосфатний кістяк, в якому цукру пов'язані один з одним ковалентними зв'язками через залишок фосфорної кислоти. Усередині спіралі знаходяться органічні азотисті основи, поєднані попарно водневими зв'язками. Сенс копіювання інформації, що зберігається в одній хромосомі, і передачі цієї інформації новій клітині без змін дуже простий: необхідно розкрутити подвійну спіраль ДНК і розірвати водневі зв'язки, які разом утримують пари азотистих основ. З цього моменту з'являються два одинарні ланцюги з азотистими основами, що виступають із сахарофосфатного кістяка. Згадайте, що кожна азотна основа одного з ланцюгів ДНК завжди утворює зв'язки з цілком певною основою іншого ланцюга ДНК. Так, аденін зв'язується лише з тиміном, а гуанін – лише з цитозином. У цьому полягає принцип комплементарності азотистих основ. Згадайте також, що нуклеотиди є молекулами, що складаються з органічної основи і фосфорильованого цукру. Теоретично, якщо необхідні нуклеотиди присутні в нуклеоплазмі, кожна азотна основа кожного ланцюга ДНК підбере собі відповідну пару. Існує специфічний та дуже важливий фермент, ДНК-полімераза, який робить цю реакцію можливою. Сахара новоствореного дочірнього ланцюга ДНК з'єднуються один з одним за допомогою фосфодіефірних зв'язків, в результаті чого утворюється сахарофосфатний кістяк - і ось воно відбулося! У вас є дві ідентичні хромосоми там, де тільки-но була одна (рис. 2.7). Звичайно, насправді процес реплікації молекули ДНК у клітині відбувається набагато складніше і залежить від функціонування цілого ряду ферментів, але про це ми ще скажемо нижче. А зараз давайте уважніше розглянемо процес поділу клітини. Спочатку пари хромосом перебувають у ядрі деспіралізованому стані і невиразні під мікроскопом. Про такі клітини кажуть, що вони перебувають у інтерфазі. Вони генетичний матеріал, відомий як хроматин, є сплутану масу і настільки компактний, наскільки тоді, коли хромосоми помітні під мікроскопом. Хроматін отримав свою назву через те, що він забарвлювався у темний колір спеціальними кольоровими барвниками. Це сталося ще задовго до того, як люди дізналися про його справжню функцію. Як згадувалося, соматичні клітини людини мають 23 пари хромосом. Двадцять дві пари є так звані аутосомні хромосоми, однакові у чоловіків та жінок. Двадцять третя пара представлена статевими хромосомами, які різні у чоловіків та жінок. Жінки мають дві великі хромосоми, названі X-хромосомами, а чоловіки – одну велику Х-хромосому та одну маленьку Y-хромосому. У хромосом однієї пари однотипна будова. Проте хромосоми, отримані від різних батьків, несуть різну інформацію, й у сенсі хромосомні пари не ідентичні.
Мітоз показаний на рис. 2.8 - універсальний спосіб поділу будь-яких еукаріотичних клітин. Мітоз складається з чотирьох основних чітко помітних фаз: профази, метафази, анафази, телофази. Коли клітина починає ділити, генетичний матеріал кожної хромосоми вже подвоївся. У профазі число хромосом відповідає 46, але кожна подвійна хромосома, тобто складається з двох сестринських хроматид.
Щоб приступити до подвоєння генетичного матеріалу, клітина має вирішити важке завдання - розкрутити спіраль ДНК. Більшість з нас думають, що хромосома не така вже й довга, тому що саме такими ми звикли бачити їх на всіх малюнках. Насправді ж в інтерфазній клітині молекула ДНК знаходиться в суперспіралізованому стані, тобто дуже щільно скручена і в певному порядку. Показовим є той факт, що якби молекула ДНК знаходилася всередині клітини в розкрученому стані, то її довжина становила б близько 1 м! Тільки уявіть собі, наскільки складним і впорядкованим має бути упаковка такої великої кількості генетичного матеріалу всередині мікроскопічної клітини. Звідси стає зрозумілим, чому в еукаріотичних клітин подвійна спіраль молекули ДНК упакована в компактну структуру, утворену за рахунок взаємодії з цілим рядом білків, званих гістонами. Таким чином, хромосомні білки упаковують гігантську молекулу в ядро об'ємом всього кілька кубічних мікрон (рис. 2.9). Безпосередньо перед початком процесу реплікації молекула ДНК розкручується ферментом хеліказою. Потім водневі зв'язки розриваються, ланцюги розходяться, і їх матриці за принципом комплементарності утворюються дочірні ланцюга. Деление клітин суворо контролюється. Існують клітини, які діляться досить часто. До них відносяться клітини кісткового мозку. Інші клітини, наприклад клітини кістки, діляться головним чином під час активного зростання органу або його ушкодження. Нарешті, існують клітини, які діляться дуже рідко чи взагалі не діляться. До такого типу належать зрілі нервові клітини. Неконтрольований поділ і зростання клітин є ознакою їхнього ракового переродження. Процес поділу клітин потребує регуляторів, які називаються факторами активації. Прикладами таких факторій є фактори зростання. Вивчення факторів зростання один з напрямків, що найбільш швидко розвиваються, на стику сучасної біології та медицини. Багато чинників нині виділено і частково охарактеризовано.
Фактори активації зазвичай працюють, змінюючи форму ДНК і дозволяючи ДНК-полімеразі, відповідальної за реплікацію ДНК, зв'язатися з нею. ДНК-полімераза спочатку знаходить специфічну ділянку в молекулі ДНК. Ця ділянка, що має вигляд петлі, необхідна для зв'язування ДНК-полімерази з ДНК. У свою чергу він утворюється при зв'язуванні фактора активації з ДНК. Ділянка молекули ДНК, де відбувається приєднання ДНК-полімерази та починається процес реплікації, називається точкою початку реплікації. Слід також знати, що для ініціації синтезу ДНК потрібні короткі послідовності РНК, що виконують функції затравок (праймерів). Зв'язавшись із молекулою ДНК, ДНК-полімераза просувається вздовж материнського ланцюга, добудовуючи дочірній ланцюг за принципом комплементарності.
Одна з найважливіших функцій ДНК-полімерази полягає у виправленні помилок у знову синтезованому ланцюзі ДНК. Дізнаючись неправильні пари основ, ДНК-полімераза повертається на один крок назад, неправильна основа видаляється з дочірнього ланцюга, а правильне вставляється. В результаті кількість помилок при реплікації ДНК становить приблизно одну помилку на 10-100 млрд пар основ. Проте, оскільки клітини діляться досить часто, помилки неминуче накопичуються. Багато з цих помилок позначаються на життєздатності клітин. Якщо клітини, в яких відбулася мутація, все ж таки виживають, імунна система руйнує їх за допомогою спеціальних, пристосованих для цих цілей клітин.
Дозвольте тепер повернутися до процесу поділу клітини. У профазі мітозу хромосоми складаються із двох хроматид, що є результатом реплікації ДНК в інтерфазі. Однак під мікроскопом їх реєструвати не вдається, тому що в профазі сестринські хроматиди тісно стикаються один з одним, взаємно спіралізуючись одна щодо іншої. У прометафазі ядерна мембрана руйнується, і хромосоми починають рухатися до екваторіальної площини клітини. Цей процес забезпечує нормальну розбіжність хромосом до полюсів клітини під час клітинного поділу, в результаті чого в кожну з клітин, що знову утворюються, потрапляє повний набір хромосом. Сестринські хроматиди однієї хромосоми пов'язані один з одним у точці, названій центроміром. В області центроміру розташований кінетохор – складна білкова структура, пов'язана з ДНК центромірної ділянки. До цієї зони під час мітозу прикріплюються нитки веретена поділу. Як ви вже спостерігали, на полюсах клітини розташовані центріолі, кількість яких перед поділом клітини також збільшується вдвічі. Центріолі беруть участь у формуванні ниток веретена поділу. Кожна нитка веретена прикріплюється до однієї хромосоми в області кінетохору. У другій фазі мітозу, метафазі, хромосоми, що складаються з двох хроматид, вишиковуються в екваторіальній площині клітини.
Нитки веретена поділу розтягують хромосоми, які з цього моменту називають дочірніми хромосомами, до протилежних кінців клітини. Фаза, протягом якої хромосоми переміщуються у різні боки, називається анафазою. Якщо все відбувається за планом, центріоль ділиться, і сестринські хроматиди розходяться до полюсів клітини. Якщо хромосоми не вишиковуються в екваторіальній площині клітини в належному порядку, процес поділу клітини може порушитися.
Після того, як хромосоми розійшлися до протилежних кінців батьківської клітини, клітина ділиться надвоє в результаті цитокінезу. Четверта та заключна стадія мітозу називається телофазою. Клітини, що розмножуються шляхом мітозу, як згадувалося, називаються соматичними. До них відносяться всі клітини нашого організму, за винятком клітин, які згодом стануть сперматозоїдами чи яйцеклітинами. Клітини, які роблять внесок у освіту нового організму, називаються статевими клітинами. Статеві клітини утворюються в результаті мейозу, який ми розглянемо в наступному розділі.
Мітоз зазвичай проходить без будь-якої затримки. Однак будь-якої миті в організмі людини відбуваються мільйони клітинних поділів. Іноді у клітинах виявляється дуже багато генетичного матеріалу, тоді як у інших може виявитися менше норми. Зазвичай такі клітини не виживають чи видаляються імунною системою.
№85 Спадкові хвороби. Класифікація спадкових хвороб.Спадкові захворювання - захворювання, виникнення та розвиток яких пов'язане з дефектами в програмному апараті клітин, що передаються у спадок через гамети. Термін використовується щодо поліетіологічних захворювань, на відміну від вужчої групи -генні хвороби.
Залежно від виду первинно уражених клітин ущелені такі групи захворювань: хвороби внаслідок мутацій у статевих клітинах - «гаметичні», тобто. власне спадкові хвороби (наприклад, фенілкетонурія, гемофілії); ці хвороби передаються у спадок; хвороби внаслідок мутацій у соматичних клітинах – «соматичні» (наприклад, пухлини, деякі хвороби імунної аутоагресії). Ці захворювання не передаються у спадок; хвороби внаслідок комбінації мутацій у статевих та соматичних клітинах (наприклад, сімейна ретинобластома).
Окремо розглядають летальні, сублетальні та гіпогенітальні захворювання: летальні хвороби призводять до загибелі під час внутрішньоутробного розвитку (наприклад, моносомії з аутосомів, гаплоїдії, більшість поліплоїдії); сублетальні захворювання призводять до загибелі індивіда до періоду статевого дозрівання (наприклад, спадкові імунодефіцити на кшталт агам-маглобулінемії швейцарського типу, синдром Луї-Бар, деякі гемофілії); гіпогенітальні захворювання поєднуються з безпліддям (наприклад, синдроми Шерешевського-Тернера, Кляйнфелтера).
Залежно від ролі спадковості та середовища виділяють чотири групи захворювань: власне спадкові хвороби (розвиваються внаслідок мутацій); екогенетичні захворювання (розвиваються за наявності мутації під впливом специфічного фактора середовища); хвороби генетичної схильності (внесок генетичних факторів очевидний); хвороби, викликані чинниками довкілля (строго кажучи, ця група хвороб не належить до спадкових хвороб, оскільки внесок генетичних чинників у їх виникнення, розвиток та результат може бути вкрай малим, сумнівним чи невідомим).