Мендел се занимава със селекцията на грах и именно на граха, научния късмет и строгостта на експериментите на Мендел дължим откриването на основните закони на наследяването: закона за еднородността на хибридите от първо поколение, закона за разделянето и законът на независимото съчетание.
Мендел експериментира с 22 сорта грах, които се различават един от друг по 7 характеристики (цвят, текстура на семената). Мендел ръководи работата си в продължение на осем години, изучава 10 000 грахови растения. Всички форми на грах, които той изследва, бяха от чисти линии; резултатите от кръстосването на такива растения едно с друго винаги са били едни и същи. Мендел представя резултатите от работата си в статия през 1865 г., която се превръща в крайъгълен камък на генетиката. Трудно е да се каже кое заслужава повече възхищение в него и работата му – строгостта на експериментите, яснотата на представянето на резултатите, перфектното познаване на експерименталния материал или познаването на работата на неговите предшественици.
Колегите и съвременниците на Мендел не успяха да оценят важността на неговите заключения. Според A.E. Гайсинович, до края на 19 век. той е цитиран само пет пъти, а само един учен - руският ботаник И.О. Шмалхаузен - оцени важността на тази работа. В началото на 20 век обаче откритите от него закони са преоткрити почти едновременно и независимо един от друг от учените К. Коренс, Е. Чермак и К. де Фрис. Значението на тези открития веднага станало очевидно за научната общност в началото на 1900-те; тяхното признаване е свързано с определени успехи в цитологията и формирането на хипотезата за ядрената наследственост.
|
Основните модели на унаследяване на черти. Човешката наследственост е изследвана в много по-малка степен от механизмите на унаследяване на признаци в растителните и животинските организми. Въпреки това вече са получени интересни данни за наследствената зависимост на много физически характеристики на човек. Съвременното ниво на генетичната наука също ни позволява да твърдим, че всички основни модели на унаследяване на черти и законите на наследствеността, установени в експерименти с растения и животни, са валидни и за хората. Например, ако млад мъж, който има гени за кафяви очи (хомозиготни за кафяви) на двете хомоложни хромозоми, се ожени за момиче със сини очи, чиито хромозоми също съдържат само гени за сини очи, тогава тяхното потомство ще наследи цвета на очите според откритите закони от основателя на генетиката Г. Мендел още през 60-те години на миналия век в опитите си с растителни хибриди на граха. Наследяването на пола при хората и повечето животни също се случва според общите закони и е свързано с разпределението на хромозомите, които имат X- и K-образна форма (полови хромозоми). Хромозомният набор на женското тяло съдържа две X хромозоми, а мъжкото тяло съдържа една X хромозома и една Y хромозома (виж фиг. 7). Броят на такива примери може да бъде безкрайно голям и няма съмнение, че „генният портрет“ на човек (генотип) до голяма степен определя много от неговите външни свойства (фенотип). По-долу са изброени някои от наследствените черти на човек, определени от доминантни и рецесивни гени (според K-Willie, 1974). Проявата на генно влияние може да се осъществи на различни етапи от онтогенезата, но повечето от фенотипните белези се определят още преди раждането. И накрая, самото проявление на гените не е фатално, а зависи от факторите на околната среда. Например, тежко наследствено заболяване, фенилкетонурия, не се развива, ако дете, съдържащо генотиповете, които причиняват това заболяване в генотипа, започне да получава определена диета веднага след раждането. По този начин генетиката има убедителни факти, доказващи съществуването както на строго наследствени черти (например кръвни нива, хемофилия, цвят на косата и очите, черти на лицето и много други), така и на черти, които до голяма степен се определят от външната среда (напр. растеж и маса на човек, силата и сръчността на неговите мускули, склонност към болести и др.). Трябва да се отбележи, че няма пряка връзка между гени и черти: развитието на една черта може да зависи от влиянието на много гени, а един ген може да повлияе на развитието на много черти. Тази картина се усложнява допълнително от постоянната корекция от външната среда. Следователно собствеността и развитието на населението винаги са тясно свързани помежду си. Невъзможно е да си представим развитието на детето без коригиращите влияния на околната среда, както е невъзможно без генетично предопределена програма за развитие, която е синтез на историческия опит на всички предишни поколения. Нека разгледаме ролята на наследствеността и околната среда в развитието на някои физически характеристики на човек в процеса на неговото пренатално и постнатално развитие. Пренатално развитие. Формирането на органите и функционалните системи на детето в процеса на ембриогенеза е под контрола на генотипа, но факторите на околната среда играят важна роля. За ембриона майчиният организъм е основната среда. Тук, в специален орган – матката – ембрионът е относително добре защитен от много вредни влияния и получава всичко необходимо за съществуването си през плацентата. Въпреки това, особено в началните етапи на развитие, много фактори, влияещи върху майчиния организъм, също влияят върху развитието на ембриона (най-значимите са: йонизиращо лъчение, заболявания, претърпели жена по време на бременност и много химикали: алкохол, никотин, антибиотици, хормонални лекарства и др.). Трябва да се отбележи, че в пренаталния период на човек има критични периоди, когато развиващият се организъм е особено чувствителен към действието на външни фактори. Има два такива периода. Първият включва началото на пренаталното развитие, първите му три седмици. По това време настъпва полагането на всички най-важни органи и неблагоприятните ефекти през този период често водят до смъртта на ембриона. През втория критичен период (от 4-та до 7-ма седмица) настъпва по-нататъшно развитие на всички органи, като вредните външни влияния през този период могат да доведат до раждането на дете с различни физически дефекти (фиг. 9). Важно е едни и същи физически вродени дефекти да бъдат свързани с увреждане на генетичните структури и с действието на неблагоприятни външни фактори в процеса на ембриогенеза. Това е добро доказателство, че средата и наследствеността са еднакво отговорни за нормалното развитие на детето. Така че към момента на раждането всички органи на човек и неговите физиологични системи, включително нервната система, обикновено се формират. Естествено възниква въпросът: наследствено ли са предопределени свойствата на нервната система, които са в основата на умствената дейност на човек, в основата на неговия ум? Има ли готови морфологични „препарати за съзнание“ по същия начин, както съществуват за долната нервна дейност, свързана с регулирането на жизнените процеси на нашето тяло? За да отговорим на тези въпроси, нека се спрем на следващия, постнатален етап от човешкото развитие. Постнатално развитие на детето. На първо място ще разгледаме основните методи, които ни позволяват да проучим специфичното значение на околната среда и наследствеността в постнаталното развитие на децата и юношите. Провеждането на подобни експерименти върху растения е просто и се състои в изолиране на две групи организми, еднакви по генотип, и поставяне на тези групи в различни условия на съществуване. Например една група растения (контролна) се отглежда при нормални условия, друга (експериментална) - в затъмнено помещение. В резултат на такива експерименти може да се заключи, че образуването на зелен хлорофил в растенията зависи не само от наследствеността, но и от факторите на околната среда (светлина), тъй като растенията, отглеждани на тъмно, няма да съдържат този пигмент. Провеждането на подобни експерименти върху хора е невъзможно както от морална и етична гледна точка, така и от биологична. В света няма двама души с еднакви генотипове. Но... има изключения от правилото. Това са еднояйчни или хомозиготни близнаци (IB), които имат не само поразителни външни прилики, но и почти идентични генни "портрети". Раждането на еднояйчни и неидентични близнаци (TW) не е необичайно; средно една от 100 бременности завършва с раждане на повече от едно дете. Близнаци е най-отличен "материал", представен на учения от самата природа, следователно самият метод се нарича близнак. Каква е същността му? Да речем, че трябва да установите кой фактор е най-отговорен за физическото развитие и растеж на детето: наследствеността или околната среда? За целта се идентифицират двойките близнаци, извършва се тяхното морфологично изследване и се изучава начина им на живот (обикновено близнаците, особено еднояйчните, имат сходни интереси и близки условия на възпитание). Въз основа на проведените изследвания се установява коефициентът на сходство (съгласуваност) между близнаците и се прави извод за ролята на наследствеността или средата в развитието на определени качества. В табл. За съжаление, учителите все още вършат малко работа за идентифициране на надарените деца, което често се свързва с подценяване на физиологичната и психическа индивидуалност на децата, с непознаване на елементарната биология на детето. Слабата работа при идентифициране на детската надареност може да се обясни и с все още ниското ниво на изследвания в тази област и липсата на точни научни критерии, по които учителят би могъл лесно да открие наследствените наклонности на детето. Съществува само тясна връзка между надареността и високото ниво на възбудимост на нервната система, чийто симптом често е рязък дисбаланс (психопатия) на детето. Учителят трябва да бъде особено внимателен към такива деца. Това е необходимо преди всичко от хигиенна страна, тъй като грешните действия на учителя могат да доведат до развитие на екстремни черти на характера в патологични. Също така е необходимо за навременното идентифициране на наследствените наклонности и тяхното оптимално развитие. Важно е да запомните, че очевидната липса на склонност към учене, външен мързел и недисциплина не са достатъчни доказателства за липса на способности. Има много доказателства от историята, че това заключение е вярно. Създателят на теорията на относителността А. Айнщайн в детството не проявява отворени способности и едва на 9-годишна възраст успява да отиде в подготвително училище. А. Хумболт (1769-1859) и Л. Пастьор (1822-1895), изключителният английски физик И. Нютон, бяха известни като мързелив и неспособен ученик в училище. Може да се предположи, че умелото отношение към тях от родители и учители в крайна сметка създава всички необходими условия за развитие на талантите. Освен това най-важното качество на гения е упоритата работа. Великият американски изобретател Т. Едисън пише: „Гениалността е един процент вдъхновение, а деветдесет и девет процента е изпотяване“. Който не се научи да се "поти", никога няма да може да постигне голям успех в каквато и да е човешка дейност, въпреки най-изявените наследствени наклонности. Кой, ако не учител, трябва преди всичко да възпита трудолюбие в детето - това е най-важният фактор за прогресивното развитие на личността, основното условие за формиране на всякакви способности. По този начин наследствеността осигурява само "суровина", а околната среда извършва основната си "преработка". Човек, който е роден дори с най-благоприятни наклонности, но живее в среда, която пречи на развитието на способностите му, ще остане посредствен, така че основната отговорност за развитието на интелектуалните способности на детето пада върху възпитателите. Независимо от това, ролята на околната среда не трябва да се абсолютизира, не трябва да се забравя, че „материалът“, доставен от наследствеността, не винаги е с високо качество и дори най-талантливият скулптор няма да може да извая скулптура от пясък, точно както мраморен блок сам по себе си няма да се превърне в произведение на изкуството... №67 - бр.68. Автозомно доминантно унаследяване- вид наследяване, при което един мутантен алел, локализиран в автозомата, е достатъчен, за да се изрази заболяването (или черта). Автозомно рецесивно унаследяване- видът на наследяване на черта или заболяване, при което мутантен алел, локализиран в автозома, трябва да бъде наследен от двамата родители. |
№69. Свързано с пола наследяване -наследяване на ген, разположен в половите хромозоми. Наследяването на черти, които се проявяват само при индивиди от същия пол, но не се определят от гени, разположени върху половите хромозоми, се нарича наследяване с ограничени полове.
Наследяването, свързано с X хромозомата, е наследяване на гени, когато мъжкият пол е хетерогаметичен и се характеризира с наличието на Y хромозома (XY), а женските са хомогаметични и имат две X хромозоми (XX). Всички бозайници (включително хората), повечето насекоми и влечуги притежават този тип наследство.
№70 Холандски тип наследство.Холандрическо наследяване (holandricinheritance) - наследяване на черта, контролирана от ген, разположен в нехомоложна част на Y-хромозомата. Холандрикгенът е ген, свързан с мъжката полова Y хромозома. Признаците, определени от Г. г, се предават само по мъжка линия – от баща на син.
№71 С X-свързан тип наследяванемутантният ген се намира на Х хромозомата. Ако мутацията има доминиращ ефект, тогава и мъжете, и жените могат да бъдат болни. Заболяването обаче се предава от болен баща със 100% вероятност само на момичета, но не и на момчета, които получават Y хромозома от баща си (фиг. 5). Вероятността за предаване на доминантна X-свързана мутация от болна майка на деца е 50%. Заболяването е еднакво вероятно да бъде наследено както от дъщерята, така и от сина.
Към доминанта,свързан с Х хромозомата, типът на унаследяване е патологията, позната на педиатрите, рахит, устойчив на витамин D (синоними: хипофосфатемия, фамилна-X-свързана хипофосфатемия, фосфатен диабет). Диагнозата на този тежък рахит, който не преминава под въздействието на големи дози витамин D, се потвърждава от наличието на подобно заболяване при някои от роднините, мъже и жени.
По-често Х-свързани заболяваниясе наследяват по рецесивен начин. Отличителна черта на заболявания с рецесивниX-свързан тип наследяване е, че мъжете са болни в семейството и те наследяват мутантния алел от своята практически здрава майка, хетерозиготна за мутантния алел. При съставянето на родословие такива майки често имат болни братя или чичовци. Болните мъже могат да предадат болестта си само през поколение и само на внуци (но не и на внучки) чрез здравата си, но хетерозиготна дъщеря.
Така, ако проследите в родословното наследство по мъжка линиярецесивно Х-свързано заболяване, получавате нещо като "ход на шах рицар".
Повечетоизвестни Х-свързани заболяваниясахемофилия А и В , както и най-тежката патология на мускулната система -Мускулна дистрофия на Дюшен Бекер ... Развитието на хемофилия А се основава на мутации в гена, отговорен за синтеза на фактор VIII на кръвосъсирването, а при хемофилия В, фактор IX на кръвосъсирването е дефектен. И двата гена, кодиращи фактори VIII и IX, са разположени на дългото рамо на X хромозомата съответно при q28 и q27.1-2. Известно е, че при хемофилия има нарушение на съсирването на кръвта и най-малките порязвания могат да доведат до смърт на пациент без специални хематологични грижи. Имайте предвид, че при жените - носители на гена на хемофилията (т.нар. "проводници"), в някои случаи има и склонност към кървене, което се изразява в обилна менструация и продължително кървене по време на раждане. Това обстоятелство трябва да се има предвид от акушер-гинеколозите при работа с жени, които носят мутантни алели в някой от гените на хемофилия А или В.
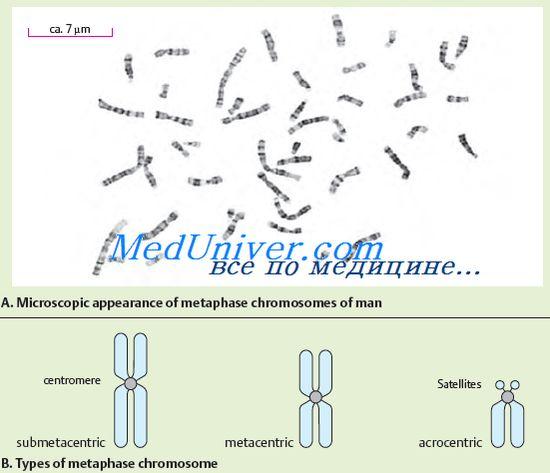
№72 Структурата и функцията на хромозомите. Хромозомната морфология се вижда най-добре в клетката на етап метафаза. Хромозомата се състои от две пръчковидни тела - хроматиди. И двете хроматиди на всяка хромозома са генетично идентични една с друга. Хромозомите са диференцирани по дължина. Хромозомите имат центромер или първично свиване, две теломери и две рамена. На някои хромозоми са изолирани вторични стеснения и сателити. Движението на хромозомата определя центромерата, която има сложна структура. Центромерната ДНК се отличава със своята характерна нуклеотидна последователност и специфични протеини. В зависимост от местоположението на центромерите се разграничават акроцентрични, субметацентрични и метацентрични хромозоми. Както бе споменато по-горе, някои хромозоми имат вторични стеснения. Те, за разлика от първичната констрикция (центромера), не служат като място за закрепване на нишките на вретеното и не играят никаква роля в движението на хромозомите. Някои вторични стеснения са свързани с образуването на нуклеоли, в този случай те се наричат нуклеоларни организатори. Ядрените организатори съдържат гените, отговорни за синтеза на РНК. Функцията на другите вторични стеснения все още не е ясна. Някои акроцентрични хромозоми имат сателити - области, които са свързани с останалата част от хромозомата чрез тънка нишка от хроматин. Формата и размерът на сателита са постоянни за дадена хромозома. При хората има сателити в пет двойки хромозоми. Краищата на хромозомите, богати на структурен хетерохроматин, се наричат теломери. Теломерите предотвратяват залепването на краищата на хромозомите след редупликация и по този начин допринасят за запазването на тяхната цялост. Следователно, теломерите са отговорни за съществуването на хромозомите като отделни единици. Хромозомите, които имат един и същ генен ред, се наричат хомоложни. Те имат една и съща структура (дължина, местоположение на центромера и т.н.). Нехомоложните хромозоми имат различен генен набор и различна структура. Изследването на фината структура на хромозомите показа, че те са съставени от ДНК, протеин и малко количество РНК. Молекулата на ДНК носи отрицателни заряди, разпределени по цялата си дължина, а прикрепените към нея протеини – хистони са положително заредени. Този ДНК-протеинов комплекс се нарича хроматин. Хроматинът може да има различна степен на кондензация. Кондензираният хроматин се нарича хетерохроматин, декондензираният хроматин се нарича еухроматин. Степента на декондензация на хроматина отразява неговото функционално състояние. Хетерохроматиновите региони са функционално по-малко активни от регионите на еухроматина, в които повечето от гените са локализирани. Разграничаване на структурен хетерохроматин, чието количество се различава в различните хромозоми, но постоянно се намира в близките центромерни области. В допълнение към структурния хетерохроматин, има незадължителен хетерохроматин, който се появява в хромозомата по време на супернавиване на еухроматични области. Потвърждение за съществуването на това явление в човешките хромозоми е фактът на генетична инактивация на една Х хромозома в соматичните клетки на жената. Същността му се крие във факта, че съществува еволюционно формиран механизъм на инактивиране на втората доза гени, локализирани в X хромозомата, в резултат на което, въпреки различния брой X хромозоми в мъжките и женските организми, броят на гените функционирането в тях е равностойно. Хроматинът е максимално кондензиран при митотично клетъчно делене, след което може да бъде открит под формата на плътни хромозоми.Размерите на ДНК молекулите на хромозомите са огромни. Всяка хромозома е представена от една ДНК молекула. Те могат да достигнат стотици микрометри и дори сантиметри. От човешките хромозоми най-голямата е първата; нейната ДНК е с обща дължина до 7 см. Общата дължина на ДНК молекулите на всички хромозоми на една човешка клетка е 170 см. Въпреки гигантския размер на ДНК молекулите, тя е доста гъсто опакована в хромозоми. Тази специфична опаковка на хромозомната ДНК се осигурява от хистонови протеини. Хистоните са подредени по дължината на ДНК молекулата под формата на блокове. Един блок съдържа 8 хистонови молекули, образуващи нуклеозома (формация, състояща се от ДНК верига, навита около хистонов октамер). Нуклеозомата е с размер около 10 nm. Нуклеозомите изглеждат като мъниста, нанизани на връв. Нуклеозомите и ДНК участъците, които ги свързват, са плътно опаковани под формата на спирала; има шест нуклеозоми за всеки завой на такава спирала. Така се формира структурата на хромозомата. Наследствената информация на организма е строго подредена според отделните хромозоми. Всеки организъм се характеризира със специфичен набор от хромозоми (брой, размер и структура), който се нарича кариотип. Човешкият кариотип е представен от двадесет и четири различни хромозоми (22 двойки автозоми, X и Y хромозоми). Кариотипът е паспортът на вида. Анализът на кариотипа може да открие нарушения, които могат да доведат до аномалии в развитието, наследствени заболявания или смърт на плода и ембрионите в ранните етапи на развитие. Дълго време се смяташе, че човешкият кариотип се състои от 48 хромозоми. Въпреки това, в началото на 1956 г. е публикуван доклад, според който броят на хромозомите в човешкия кариотип е 46. Човешките хромозоми се различават по размер, местоположение на центромерите и вторичните стеснения. За първи път разделянето на кариотипа на групи е извършено през 1960 г. на конференция в Денвър (САЩ). Описанието на човешкия кариотип първоначално включва следните два принципа: разположението на хромозомите по дължината им; групиране на хромозоми по местоположение на центромера (метацентрично, субметацентрично, акроцентрично). Точното постоянство на броя на хромозомите, тяхната индивидуалност и сложността на структурата показват важността на функцията, която изпълняват. Хромозомите изпълняват функцията на основния генетичен апарат на клетката. При тях гените са подредени в линеен ред, всеки от които заема строго определено място (локус) в хромозомата. Всяка хромозома съдържа много гени, но за нормалното развитие на организма е необходим набор от гени от пълен набор от хромозоми.
№73. Наследствеността се основава на способността на всички живи организми да натрупват, съхраняват и предават наследствена информация на потомството. Тази една от най-важните качествени характеристики на живата материя е свързана с нуклеиновите киселини – дезоксирибонуклеинова (ДНК) и рибонуклеинова (РНК). Водещата роля принадлежи на ДНК - най-дългата молекула на живите организми, концентрирана в ядрата на клетките и представляваща нейния наследствен апарат. Голямата дължина на молекулата на ДНК дава възможност да се „запишат” върху нея, като на телеграфна лента, всички основни свойства на бъдещия организъм и програмата за неговото развитие. Такъв "запис" се извършва с помощта на специален "нуклеинов език", или "нуклеинов код", чиято същност е да промени реда на четирите химични съединения, които изграждат ДНК. Образно казано, "нуклеиновият език" се състои от четири букви, от които се изграждат отделни думи и цели изречения на „нуклеиновия език“.
На такава нуклеинова "ивица" могат да се разграничат отделни независими области, включително описание на програмата за развитие на една черта. Те се наричат гени. Всяка молекула ДНК включва стотици гени и е програма за развитие на много характеристики и свойства на организма. Комбинирайки се със специални протеини и някои други вещества, молекулите на ДНК образуват специални образувания в ядрото - така наречените хромозоми.
Организация на генетичния материал. Гени [от гръцки. genos, раждане] - единица за наследственост, част от ДНК, която заема определено място в хромозомата. От гледна точка на генетиката, генът е наследствен фактор и неделима единица от генетичен материал. Структурният ген (цистрон) е ДНК фрагмент, участващ в образуването на полипептидна верига. Състои се от водеща последователност, кодиращи фрагменти (екзони), последователности на вмъкване (нитрони) и крайна последователност. Тъй като някои протеини се състоят от повече от една субединица, формулировката "един ген - един ензим" във връзка с хетеромултимерен (тоест състоящ се от две или повече различни полипептидни субединици) протеин трябва да се тълкува като "един ген - една полипептидна верига" .
Генотипът е съвкупност от гени в организма. Още в древни времена хората емпирично са използвали законите за наследяване. На базата на този опит се е развила селекция [от лат. selectio, select] - науката за методите за създаване на нови сортове растения и породи животни чрез селекция и кръстосване. Доскоро генотипът изглеждаше непревземаем, неподвластен на човешките действия. Откриването на структурата на гените направи възможно те да се изолират в изолирана форма, да се синтезират биохимично и дори да се въведат в тялото. Стана възможно да се повлияе на гена, без да се отделя от тялото. Всичко това създаде предпоставки за манипулиране на генотипа.
№74 Структурата на нуклеиновите киселини.
Нуклеинова киселина(от лат. nucleus - ядро) - високомолекулно органично съединение, биополимер (полинуклеотид), образуван от остатъци от нуклеотиди. Нуклеиновите киселини ДНК и РНК присъстват в клетките на всички живи организми и изпълняват най-важните функции за съхранение, предаване и реализиране на наследствена информация. Полимерните форми на нуклеиновите киселини се наричат полинуклеотиди. Веригите от нуклеотиди са свързани чрез остатък от фосфорна киселина (фосфодиестерна връзка). Тъй като в нуклеотидите има само два вида хетероциклични молекули, рибоза и дезоксирибоза, има само два вида нуклеинови киселини - дезоксирибонуклеинова киселина (ДНК) и рибонуклеинова киселина (РНК).
Мономерните форми също се намират в клетките и играят важна роля в сигнализирането или съхранението на енергия. Най-известният РНК мономер е АТФ, аденозинтрифосфорната киселина, най-важният акумулатор на енергия в клетката.
Една от основните функции на нуклеиновите киселини е да определят протеиновия синтез. Информацията за структурата на протеините, кодирани в нуклеотидната последователност на ДНК, трябва да се предава от едно поколение на друго и следователно трябва да бъде правилно копирана, т.е. синтез на абсолютно същата ДНК молекула (репликация).
№75 RepДНК ликиране процесът на синтеза на дъщерна молекула на дезоксирибонуклеинова киселина върху матрицата на родителската ДНК молекула. По време на последващото делене на клетката майка, всяка дъщерна клетка получава едно копие на ДНК молекулата, която е идентична с ДНК на оригиналната клетка майка. Този процес осигурява точното предаване на генетична информация от поколение на поколение. Репликацията на ДНК се осъществява от сложен ензимен комплекс, състоящ се от 15-20 различни протеина, наречен реписома. Репликацията на ДНК е ключово събитие в клетъчното делене. Важно е по време на разделянето ДНК да се репликира напълно и само веднъж. Това се осигурява от определени механизми за регулиране на репликацията на ДНК. Репликацията се извършва на три етапа:
иницииране на репликация
удължаване
прекратяване на репликацията.
Регулирането на репликацията се извършва главно в началния етап. Това е доста лесно за изпълнение, тъй като репликацията може да започне не от която и да е част от ДНК, а от строго определена, наречена място на иницииране на репликация. В генома може да има само едно или много такива места. Концепцията за репликон е тясно свързана с концепцията за място за иницииране на репликация. Репликонът е участък от ДНК, който съдържа място за иницииране на репликация и се репликира, след като синтезът на ДНК започне от това място. Геномите на бактериите, като правило, представляват един репликон, което означава, че репликацията на целия геном е резултат само от един акт на иницииране на репликация. Геномите на еукариотите (както и техните индивидуални хромозоми) се състоят от голям брой независими репликони, което значително намалява общото време на репликация на отделна хромозома. Молекулните механизми, които контролират броя на действията за иницииране на репликация във всяко място по време на един цикъл на клетъчно делене, се наричат контрол на броя на копията. В допълнение към хромозомната ДНК, бактериалните клетки често съдържат плазмиди, които са индивидуални репликони. Плазмидите имат свои собствени механизми за контрол на копирането: те могат да осигурят синтеза само на едно копие на плазмида на клетъчен цикъл или хиляди копия.
Репликацията започва от мястото на иницииране на репликацията с развиване на двойната спирала на ДНК, с образуването на репликационна вилка – мястото на директна репликация на ДНК. Всеки сайт може да образува една или две разклонения за репликация, в зависимост от това дали репликацията е еднопосочна или двупосочна. Двупосочната репликация е по-често срещана. Известно време след началото на репликацията в електронен микроскоп е възможно да се наблюдава репликационното око - участък от хромозомата, където ДНК вече е репликирана, заобиколен от по-разширени участъци от нерепликирана ДНК.
В репликационната вилица ДНК копира голям протеинов комплекс (репликазома), чийто ключов ензим е ДНК полимераза. Репликационната вилка се движи със скорост от около 100 000 базови двойки в минута при прокариотите и 500-5 000 при еукариотите.
№76 Структурата и видовете хромозоми.Ядрото на всяка соматична клетка на човешкото тяло съдържа46 хромозоми... комплектхромозоми всеки индивид, както нормален, така и патологичен, се нарича кариотип.
От46 хромозомикоито съставляват човешкия хромозомен набор, 44 или 22 двойки представляват автозомнихромозоми , последната двойка са полови хромозоми. При жените структурата на половите хромозоми обикновено е представена от две X хромозоми, а при мъжете - от X и Y хромозоми.
Във всичкодвойки хромозомикакто автозомно, така и полов, едната от хромозомите се получава от бащата, а другата от майката. Хромозомите от една двойка се наричат хомолози или хомоложни хромозоми. Репродуктивните клетки (сперма и яйцеклетки) съдържат хаплоиден набор от хромозоми, т.е. 23 хромозоми.
 Сперматозоидите се делят на два вида в зависимост от това дали съдържатхромозомаX или Y. Всички яйца обикновено съдържатсамо хромозома
Х.
Сперматозоидите се делят на два вида в зависимост от това дали съдържатхромозомаX или Y. Всички яйца обикновено съдържатсамо хромозома
Х.
хромозомиса ясно видими след специално оцветяване по време на клетъчното делене, когато хромозомите са максимално спирализирани. В този случай във всяка хромозома се открива стеснение, което се нарича центромер. Центромерата разделя хромозомата на късо рамо (означено с буквата "p") и дълго рамо (означено с буквата "q"). Центромерът определя движението на хромозомата по време на клетъчното делене. Според позицията на центромера хромозомите се класифицират в няколко групи. Ако центромерата е разположена в средата на хромозомата, тогава такава хромозома се нарича метацентрична, ако центромерата е разположена по-близо до един от краищата на хромозомата, тогава тя се нарича акроцентрична. Някои акроцентрични хромозоми имат така наречените сателити, които образуват нуклеоли в неделяща се клетка. Ядрата съдържат множество копия на рРНК. Освен това се разграничават субметацентрични хромозоми, когато центромерът е разположен не в средата на хромозомата, а донякъде изместен към един от краищата, но не толкова значително, колкото при акроцентричните хромозоми.
Краищата на всекираменна хромозоманаречени теломери. Установено е, че теломерите играят важна роля в поддържането на стабилността на хромозомите. Теломерите съдържат голям брой повторения на TTAGGG нуклеотидната последователност, така наречените тандемни повторения. Обикновено по време на клетъчното делене има намаляване на броя на тези повторения теломери .
Въпреки това, всеки път те се завършват с помощта на специален ензим, наречентеломераза... Намаляването на активността на този ензим води до скъсяване на теломерите, което се смята за причина за клетъчната смърт и обикновено съпътства стареенето.
№77. Класификация на хромозомите в Денвър.Класификацията и номенклатурата на равномерно оцветените човешки хромозоми са приети за първи път на международна среща през 1960 г. в Денвър, по-късно донякъде модифицирани и допълнени (Лондон, 1963 г. и Чикаго, 1966 г.). Според класификацията на Денвър всички човешки хромозоми са разделени на 7 групи, подредени в низходящ ред на тяхната дължина и като се вземе предвид индексът на центриола (съотношението на дължината на късото рамо към дължината на цялата хромозома, изразено като процент). Групите се обозначават с букви на английската азбука от A до G. Всички двойки хромозоми обикновено се номерират с арабски цифри. Характеристиките на групите са представени в табл. 4. Предложената класификация позволи ясно да се разграничат хромозомите, принадлежащи към различни групи. От 1960 г. започва бързото развитие на клиничната цитогенетика: през 1959 г. J. Lejeune открива хромозомната природа на синдрома на Даун; K. Ford, P. Jacobs и J. Strong описват особеностите на кариотипа при синдромите на Klinefelter и Turner; в началото на 70-те години. хромозомната природа на синдромите на Едуардс и Патау, синдромът на "котешки плач" е открит; описва хромозомна нестабилност при редица наследствени синдроми и злокачествени заболявания. В същото време прилагането на метода за получаване на равномерно оцветени хромозоми се оказа недостатъчно ефективно за идентифициране на хромозоми. Недостатъкът на класификацията в Денвър е, че диференциацията на хомоложни двойки в рамките на група хромозоми често среща непреодолими трудности.
Таблица 4^ Групи хромозоми и техните характеристики
|
Група |
Номер на хромозомата |
Местоположение Centromere |
Индекс на центриола (%) |
Забележка |
|
Най-големият метацентричен |
На дългото рамо може да има вторично свиване |
|||
|
Най-големият субметацентричен | ||||
|
Голям метацентричен |
20% по-къс от първия |
|||
|
Голям субметацентричен | ||||
|
6-12 и Х хромозома |
Средно субметацентричен |
На 9-ти често има вторично свиване |
||
|
Средно акроцентричен |
На всички вторични стеснения |
|||
|
Малък метацентричен |
Вторичното свиване се появява в 10% от случаите. |
|||
|
Малък субметацентричен | ||||
|
Най-малкият метацентричен | ||||
|
21-22 и Y-хромозома |
Най-малкият акроцентрик |
На 21-ви и 22-ри вторични стеснения |
№78 Биосинтеза на протеин в клетката.Синтезът на протеин е много сложен, многоетапен процес. Провежда се в специални органели - рибозоми. Клетката съдържа голям брой рибозоми. Например, E. coli има около 20 000 от тях.
Как протича протеиновият синтез в рибозомите?
Протеиновите молекули са по същество полипептидни вериги, съставени от отделни аминокиселини. Но аминокиселините не са достатъчно активни, за да се свързват сами. Следователно, преди да се свържат помежду си и да образуват протеинова молекула, аминокиселините трябва да бъдат активирани. Това активиране става под действието на специални ензими. Освен това всяка аминокиселина има свой собствен ензим, специално настроен към нея.
Източникът на енергия за това (както и за много процеси в клетката) е аденозин трифосфат (АТФ).
В резултат на активирането аминокиселината става по-лабилна и под действието на същия ензим се свързва с t-RNA.
Важно е всяка аминокиселина да съответства на строго специфична t-RNA. Тя намира "своята" аминокиселина и я прехвърля в рибозомата. Следователно тази РНК се нарича транспортна РНК.
Следователно, различни активирани аминокиселини, комбинирани с техните t-RNA, влизат в рибозомата. Рибозомата е като че ли транспортна лента за сглобяване на протеинова верига от различни аминокиселини, влизащи в нея.
Оказва се, че едновременно с t-РНК, върху която "седи" собствената й аминокиселина, рибозомата получава "сигнал" от ДНК, съдържаща се в ядрото. В съответствие с този сигнал в рибозомата се синтезира един или друг протеин, един или друг ензим (тъй като ензимите са протеини).
Насочващото влияние на ДНК върху протеиновия синтез не се осъществява директно, а с помощта на специален медиатор, тази форма на РНК, която се нарича информационна или информационна РНК (m-RNA или i-RNA).
Информационната РНК се синтезира в ядрото от йод под въздействието на ДНК, поради което съставът й отразява състава на ДНК. Молекулата на РНК е като матрица под формата на ДНК.
Синтезираната m-RNA влиза в рибозомата и като че ли предава на тази структура план – в какъв ред трябва да се комбинират помежду си активираните аминокиселини, влизащи в рибозомата, за да може да се синтезира определен протеин. В противен случай генетичната информация, кодирана в ДНК, се прехвърля към m-RNA и след това към протеина.
№79 мутация (лат.mutatio - промяна) -устойчива (т.е. такава, която може да бъде наследена от потомците на дадена клетка или организъм) промяна в генотипа, която настъпва под влияние на външната или вътрешната среда. Терминът е въведен от Хуго де Фрис. Процесът на възникване на мутации се нарича мутагенеза.
Геномна: - полиплоидизация (образуване на организми или клетки, чийто геном е представен от повече от два (3n, 4n, 6n и др.) набора хромозоми) и неуплоидия (хетероплоидия) - промяна в броя на хромозомите, които не е кратно на хаплоидния набор (виж Inge-Vechtomov, 1989). В зависимост от произхода на хромозомните набори, полиплоидите разграничават алополиплоиди, които имат набори от хромозоми, получени чрез хибридизация от различни видове, и автополиплоиди, които имат увеличение на броя на наборите от хромозоми на собствения си геном, кратно на n.
При хромозомни мутации настъпват големи промени в структурата на отделните хромозоми. В този случай има загуба (изтриване) или удвояване на част (дублиране) от генетичния материал на една или няколко хромозоми, промяна в ориентацията на хромозомните сегменти в отделните хромозоми (инверсия), както и прехвърляне на част от генетичния материал от една хромозома в друга (транслокация) (краен случай - обединяване на цели хромозоми, т.нар. Робъртсонова транслокация, която е преходен вариант от хромозомна към геномна мутация).
На генно ниво промените в първичната структура на генната ДНК под влияние на мутации са по-малко значими, отколкото при хромозомни мутации, но генните мутации са по-чести. В резултат на генни мутации, замествания, делеции и инсерции на един или повече нуклеотиди възникват транслокации, дублирани и инверсии на различни части на гена. В случай, че само един нуклеотид се променя под действието на мутация, се говори за точкови мутации. Тъй като ДНК съдържа само два вида азотни бази - пурини и пиримидини, всички точкови мутации с замествания на основи са разделени на два класа: преходи (замяна на пурин с пурин или пиримидин с пиримидин) и трансверсия (замяна на пурин с пиримидин или обратно) . Има четири възможни генетични последици от точковите мутации: 1) запазване на значението на кодона поради дегенерацията на генетичния код (синонимно терминиране (безсмислена мутация). В генетичния код има три безсмислени кодона: кехлибар - UAG, ocp - UAA и опал - UGA (в съответствие с това името и мутациите, които водят до образуването на безсмислени триплети - например кехлибарена мутация), 4) обратно заместване (стоп кодон със сензорен кодон).
Според влиянието им върху генната експресия, мутациите се разделят на две категории: мутации от типа замествания на базовата двойка и типа на изместване на рамката (frameshift). Последните представляват делеции или вмъквания на нуклеотиди, чийто брой не е кратен на три, което е свързано с триплетната природа на генетичния код.
Първичната мутация понякога се нарича директна мутация, а мутация, която възстановява оригиналната структура на гена, се нарича обратна мутация или реверсия. Връщането към първоначалния фенотип на мутантния организъм поради възстановяването на функцията на мутантния ген често се случва не поради истинска реверсия, а поради мутация в друга част от същия ген или дори друг неалелен ген. В този случай повтарящата се мутация се нарича супресорна мутация. Генетичните механизми, чрез които мутантният фенотип се потиска, са много разнообразни.
Бъбречните мутации (спори) са устойчиви соматични мутации, възникващи в клетките на точките на растеж на растенията. Води до клонова вариация. Те се запазват по време на вегетативно размножаване. Много сортове са бъбречни мутации.
№80. Опаковане на ДНК в хромозома.Хромозомите са съставени от хроматин - смес от ДНК и протеини в съотношение ~1:1. Благодарение на взаимодействието с протеините молекулата на ДНК има компактна структура. Средната дължина на една верига от човешка ДНК (ако е разтегната) е 4 см. Тоест, ядрото на всяка клетка съдържа около 1 метър ДНК. Опаковането на ДНК молекула в хромозома има три нива (виж фигурата): 1. нуклеозоми, нуклеозомна верига 2. соленоидна спирала 3. бримки Двойна ДНК спирала, позната на всеки от нейните типове, се "навива" около комплекс от 4 двойки от белтъчни молекули образува "мънисто" - нуклеозома. Нуклеозомите, свързани помежду си от участъци от молекулата на ДНК, образуват нишка, която от своя страна е усукана под формата на соленоид (това е според един от съществуващите модели), един завой - 6 нуклеозоми. Тази структура образува бримки на следващото ниво на опаковката. В тази форма ДНК присъства в клетъчното ядро на етапа на клетъчния растеж, синтеза на неговите компоненти, включително дублирането на ДНК. Въпреки това, на различни етапи от живота на клетката, кондензацията на хроматин е различна. По време на клетъчния растеж хроматинът в ядрото не се кондензира, компактността на местоположението му е различна за различните части на хромозомата. Когато настъпи процесът на клетъчно делене, хроматинът става все по-плътен и бримките, които бяха споменати по-горе, образуват "розетки", плътно опаковани една в друга (също според един от съществуващите модели), по 18 бримки на пръстен. В резултат на това хроматинът приема формата на кондензирани хромозоми, така добре познати от учебниците.
№81. Гаметогенеза.(от Гамета и гръцки genesis - произход) процесът на развитие и образуване на полови клетки - гамети (Виж Гамети). G. мъжки гамети (сперматозои, сперма) се наричат сперматогенеза, женски гамети (ооцити) - оогенеза. При животните и растенията G. протича различно, в зависимост от мястото на мейозата в жизнения цикъл на тези организми.
При многоклетъчни животни G. се среща в специални органи - половите жлези или гонади (яйчници, тестиси и хермафродитни гонади) и се състои от три основни етапа: 1) възпроизвеждане на първичните зародишни клетки - гаметогония (сперматогония и оогония) чрез серия от последователни митози 2) растежът и съзряването на тези клетки сега се наричат гаметоцити (сперматоцити и ооцити), които, подобно на гаметогонията, имат пълен (предимно диплоиден) набор от хромозоми (вижте Хромозоми). По това време настъпва основното събитие на G. при животните - разделянето на гаметоцитите чрез мейоза, което води до намаляване (намаляване наполовина) на броя на хромозомите в тези клетки и превръщането им в хаплоидни клетки (виж Хаплоид) - сперматиди и яйцеклетки ; 3) образуването на сперматозоиди (или сперматозоиди) и яйцеклетки; в този случай яйцеклетките са облечени с редица ембрионални мембрани, а сперматозоидите придобиват флагели, които осигуряват тяхната подвижност. При женски от много животински видове мейозата и образуването на яйцеклетки завършват след проникването на спермата в цитоплазмата на ооцита, но преди сливането на ядрата на спермата и яйцеклетката.
При растенията G. се отделя от мейозата и започва в хаплоидни клетки, в спори (при висши растения, микроспори и мегаспори). От спорите се развива половото поколение на растението - хаплоидният гаметофит, в гениталиите на който - гаметангии (мъжки - антеридии (виж Anteridium), женски - архегония (виж Archegonia)) чрез митоза, G. Изключение правят голосеменните и покритосеменни растения, при които сперматогенезата върви директно в покълналата микроспора - поленова клетка. При всички нисши и висши спорови растения G. в антеридиите е множествено делене на клетките, в резултат на което се образуват голям брой дребни подвижни сперматозоиди. G. при архегония - образуване на един, два или повече ооцита. При голосеменните и покритосеменните растения мъжкият G. се състои от разделяне (чрез митоза) на ядрото на поленова клетка на генеративна и вегетативна и по-нататъшно разделяне (също чрез митоза) на генеративното ядро на два сперматозоида. Това разделяне се извършва в тръбата за поникване на цветен прашец. Женски G. в покритосеменните растения е изолирането чрез митоза на една яйцеклетка в осемядрена ембрионална торбичка. Основната разлика на G. при животните и растенията: при животните съчетава трансформацията на клетките от диплоидни в хаплоидни и образуването на хаплоидни гамети; при растенията G. се свежда до образуването на гамети от хаплоидни клетки.
№82 кариотип -набор от характеристики (брой, размер, форма и т.н.) на пълен набор от хромозоми, присъщи на клетките на даден биологичен вид (видов кариотип), на даден организъм (индивидуален кариотип) или линия (клон) от клетки. Визуалното представяне на пълен хромозомен набор (кариограма) понякога се нарича още кариотип. Появата на хромозомите се променя значително по време на клетъчния цикъл: по време на интерфазата хромозомите са локализирани в ядрото, като правило, деспирализирани и трудни за наблюдение, следователно клетките в един от етапите на тяхното делене - метафаза на митоза се използват за определяне на кариотип. За процедурата за определяне на кариотипа може да се използва всяка популация от делящи се клетки. За определяне на човешкия кариотип по правило се използват лимфоцити от периферна кръв, чийто преход от стадий на покой G0 към пролиферация се провокира от добавянето на митогенафитохемаглутинин. За определяне на кариотипа могат да се използват и клетки от костен мозък или първична култура на кожни фибробласти. За да се увеличи броят на клетките на етап метафаза, колхицинилинокадазол се добавя към клетъчната култура малко преди фиксирането, които блокират образуването на микротубули, като по този начин предотвратяват разминаването на хроматидите към полюсите на клетъчното делене и завършването на митозата.
След фиксиране препарати от метафазни хромозоми се оцветяват и фотографират; от микрографии се формира така нареченият систематичен кариотип - номериран набор от двойки хомоложни хромозоми, изображенията на хромозомите са ориентирани вертикално с къси рамена нагоре, номерирането им се извършва в намаляващ ред на размера, двойка полови хромозоми се поставя на края на комплекта (виж фиг. 1).
Исторически, първите неподробни кариотипове, които направиха възможно класифицирането според морфологията на хромозомите, бяха получени чрез оцветяване по Романовски-Гимза, но по-нататъшното детайлизиране на структурата на хромозомите в кариотипите стана възможно с появата на методи за диференциално оцветяване на хромозоми. Най-често използваната техника в медицинската генетика е оцветяването с G-диференциална хромозома.
Сравнението на комплексите от напречни белези в класическата кариотипия или региони със специфични спектрални характеристики дава възможност да се идентифицират както хомоложни хромозоми, така и техните отделни области, което дава възможност да се определят подробно хромозомни аберации - вътрешно- и междухромозомни пренареждания, придружени от нарушение от порядъка на хромозомните фрагменти (делеции, дупликации, инверсии, транслокации). Такъв анализ е от голямо значение в медицинската практика, позволявайки диагностицирането на редица хромозомни заболявания, причинени както от груби нарушения на кариотипите (аномалии в броя на хромозомите), така и от нарушение на хромозомната структура или множество клетъчни кариотипове в организма. (мозаицизъм).
№83 Мейоза- редукционно деление, деление на съзряване, метод на клетъчно делене, в резултат на което има намаляване (намаляване) на броя на хромозомите наполовина и една диплоидна клетка (съдържаща два комплекта хромозоми) след две бързо следващи една след друга деления води до 4 хаплоидни (съдържащи един набор от хромозоми).
Биологична стойност М ... е да се поддържа последователностКариотипи в редица поколения организми от този тип и осигуряване на възможност за рекомбинация) на хромозоми и гени по време на половия процес. М. е един от ключовите механизми на наследствеността
Мейозата се състои от 2 последователни деления с къса интерфаза между тях.
Профаза I - профазата на първото деление е много сложна и се състои от 5 етапа:
Лептотен или лептонема - пакетиране на хромозоми, кондензация на ДНК с образуване на хромозоми под формата на тънки нишки (хромозомите се съкращават).
Зиготена или зигонема - възниква конюгация - комбинацията от хомоложни хромозоми с образуването на структури, състоящи се от две свързани хромозоми, наречени тетради или биваленти, и тяхното по-нататъшно компактизиране.
Пахитена или пахинема - (най-дългият етап) - на някои места хомоложните хромозоми са тясно свързани, образувайки хиазми. В тях има пресичане- обмен на места между хомоложни хромозоми.
Diplotene или Diploneme - настъпва частична декондензация на хромозоми, докато част от генома може да работи, протичат процеси на транскрипция (образуване на РНК), транслация (протеинов синтез); хомоложните хромозоми остават свързани помежду си. При някои животни хромозомите в ооцитите на този етап от мейотична профаза придобиват характерна форма хромозоми с четка за лампа.
Диакинеза – ДНК се кондензира отново, доколкото е възможно, синтетичните процеси спират, ядрената обвивка се разтваря; центриолите се разминават към полюсите; хомоложните хромозоми остават свързани помежду си.
Към края на профаза I центриолите мигрират към полюсите на клетката, образуват се филаменти на вретеното на делене, ядрената мембрана и нуклеоли се разрушават
Метафаза I - двувалентните хромозоми се подреждат по екватора на клетката.
Анафаза I - микротубулите се свиват, бивалентите се разделят и хромозомите се разминават към полюсите. Важно е да се отбележи, че поради конюгирането на хромозомите в зиготена, цели хромозоми се разминават към полюсите, всяка от които се състои от две хроматиди, а не отделни хроматиди, както в митоза.
Телофаза I - хромозомите се деспирализират и се появява ядрена обвивка.
Второто деление на мейозата веднага следва първото, без изразена интерфаза: S-периодът отсъства, тъй като няма репликация на ДНК преди второто деление.
Профаза II - настъпва кондензация на хромозоми, клетъчният център се разделя и продуктите от неговото делене се отклоняват към полюсите на ядрото, ядрената обвивка се разрушава, образува се вретено на делене.
Метафаза II - едновалентните хромозоми (всяка от две хроматиди) са разположени на "екватора" (на еднакво разстояние от "полюсите" на ядрото) в една равнина, образувайки така наречената метафазна плоча.
Анафаза II - едновалентно разделяне ихроматидисе разминават към полюсите.
Телофаза II - хромозомите се деспирализират и се появява ядрена обвивка.
В резултат на това от единдиплоидна клеткачетирихаплоидни клетки... В случаите, когато мейозата е свързана сгаметогенеза(например при многоклетъчни животни), по време на развитиетоооцитипървото и второто разделяне на мейозата са рязко неравномерни. В резултат на това едно хаплоидно яйце и три т.нар редукционни тела(абортивни производни на първа и втора дивизия).
№84 Деление на клетките - митоза
Ако наблюдавате клетъчното делене под микроскоп, ще видите следното. Първо, материалът вътре в сърцевината става много плътен. Можете дори да правите разлика между дискретни структури - хромозоми. Знаете, че човешките соматични (не-полови) клетки имат 23 двойки хромозоми - по една хромозома от всяка двойка от всеки родител. Общо в човешките соматични клетки има 46 хромозоми. Всички 46 се подреждат в екваториалната равнина на клетката. На този етап е ясно видимо, че всяка от 46-те хромозоми всъщност се състои от две хроматиди, свързани една с друга в регион, подобен на бутон. Тази област е наречена центромер. На противоположните краища на клетката има две плътни петна (центриоли), всяко от които образува нишките на делителното вретено. Всяка нишка се присъединява към хромозомата в центромера. Към всяка двойка хромозоми са прикрепени две нишки - по една от всеки центриол. Тогава ще видим, че центриолите, като намотки, навиват нишките на вретено на делене, в резултат на което хромозомите се разминават към полюсите на клетката. След това съдържанието на клетката се разделя. Това е невероятна гледка.
Събитията се развиват по следния сценарий. Необходимо е да се разбере, че за клетъчното делене трябва да се реши много важна задача - трябва да се случи дублиране (репликация) на ДНК, защото именно в ДНК е кодирана цялата информация какво и как трябва да се случи в клетката. Не забравяйте, че ДНК е двойна спирала. Извън спиралата е захарно-фосфатният гръбнак, в който захарите са свързани една с друга чрез ковалентни връзки чрез остатъка от фосфорна киселина. Вътре в спиралата има органични азотни основи, свързани по двойки чрез водородни връзки. Значението на копирането на информация, съхранявана в една хромозома, и прехвърлянето на тази информация в нова клетка без промени е много просто: необходимо е да се развие двойната спирала на ДНК и да се разкъсат водородните връзки, които държат двойките азотни бази заедно. От този момент се появяват две единични вериги с азотни основи, излизащи от захарно-фосфатния гръбнак. Не забравяйте, че всяка азотна база на една от ДНК нишките винаги образува връзки с много специфична база на другата ДНК верига. И така, аденинът се свързва само с тимин, а гуанинът само с цитозин. Това е принципът на комплементарност на азотните основи. Не забравяйте също, че нуклеотидите са молекули, съставени от органична основа и фосфорилирана захар. На теория, ако необходимите нуклеотиди присъстват в нуклеоплазмата, всяка азотна база на всяка ДНК верига ще съвпадне с подходяща двойка. Има специфичен и много важен ензим, ДНК полимераза, който прави тази реакция възможна. Захарите на новообразуваната дъщерна ДНК верига са свързани помежду си чрез фосфодиестерни връзки, в резултат на което се образува захарно-фосфатен гръбнак – и така се случи! Имате две еднакви хромозоми, където току-що сте имали една (Фигура 2.7). Разбира се, всъщност процесът на репликация на ДНК молекула в клетката е много по-сложен и зависи от функционирането на редица ензими, но за това ще говорим по-долу. Сега нека разгледаме по-отблизо процеса на клетъчно делене. Първоначално двойките хромозоми са в деспирализирано състояние в ядрото и са неразличими под микроскоп. За такива клетки се казва, че са в интерфаза. При тях генетичният материал, известен като хроматин, е заплетена маса и не е толкова компактна, колкото когато хромозомите са различими под микроскоп. Хроматинът получи името си от факта, че е оцветен в тъмно със специални цветни багрила. Това се случи много преди хората да разберат за истинската му функция. Както вече споменахме, човешките соматични клетки имат 23 двойки хромозоми. Двадесет и две двойки се наричат автозомни хромозоми, които са еднакви при мъжете и жените. Двадесет и третата двойка е представена от полови хромозоми, които са различни при мъжете и жените. Жените имат две големи хромозоми, наречени X хромозоми, докато мъжете имат една голяма X хромозома и една малка Y хромозома. Хромозомите на една двойка имат същата структура. Въпреки това, хромозомите, получени от различни родители, носят различна информация и в този смисъл хромозомните двойки не са идентични.
Митозата, показана на фиг. 2.8, е универсален начин за разделяне на всякакви еукариотни клетки. Митозата се състои от четири основни отделни фази: профаза, метафаза, анафаза, телофаза. Когато една клетка започне да се дели, генетичният материал на всяка хромозома вече се е удвоил. В профаза броят на хромозомите съответства на 46, но всяка хромозома е двойна, тоест се състои от две сестрински хроматиди.
За да започне удвояване на генетичния материал, клетката трябва да реши трудна задача - да развие спиралата на ДНК. Повечето от нас смятат, че хромозомата не е толкова дълга, защото така сме свикнали да ги виждаме във всички рисунки. В действителност, в интерфазна клетка, молекулата на ДНК е в свръхнавито състояние, тоест тя е усукана много плътно и в определен ред. Показателен е фактът, че ако ДНК молекулата се намираше вътре в клетката в неусукано състояние, то дължината й би била около 1 m! Само си представете колко сложно и подредено трябва да бъде опаковането на толкова много генетичен материал в една микроскопична клетка. Следователно става ясно защо в еукариотните клетки двойната спирала на ДНК молекулата е опакована в компактна структура, образувана от взаимодействие с редица протеини, наречени хистони. Така хромозомните протеини опаковат гигантската молекула в ядро с обем само няколко кубични микрона (фиг. 2.9).Непосредствено преди началото на репликационния процес, молекулата на ДНК се развива от ензима хеликаза. Тогава водородните връзки се разкъсват, веригите се разминават и върху матрицата им се образуват дъщерни вериги по принципа на комплементарността.Клетъчното делене се контролира строго. Има клетки, които се делят често. Те включват клетки от костен мозък. Други клетки, като костните клетки, се делят главно по време на активен растеж или увреждане на органите. И накрая, има клетки, които се делят много рядко или изобщо не се делят. Този тип включва зрели нервни клетки. Неконтролираното делене и растеж на клетките е признак за тяхната ракова дегенерация. Процесът на клетъчно делене изисква регулатори, наречени фактори на активиране. Факторите на растеж са примери за такива фабрики. Изучаването на растежните фактори е една от най-бързо развиващите се области на пресечната точка на съвременната биология и медицина. Много фактори в момента са подчертани и частично характеризирани.
Факторите на активиране обикновено действат, като променят формата на ДНК и позволяват на ДНК полимеразата, която е отговорна за репликацията на ДНК, да се свърже с нея. ДНК полимеразата първо открива специфично място в ДНК молекулата. Този участък с форма на бримка е необходим за свързването на ДНК полимеразата с ДНК. От своя страна се образува, когато факторът на активиране се свърже с ДНК. Мястото на ДНК молекулата, където е прикрепена ДНК полимеразата и започва процесът на репликация, се нарича точка на произход на репликацията. Трябва също да сте наясно, че са необходими къси РНК последователности за иницииране на синтеза на ДНК, които функционират като „праймери“. Свързвайки се с ДНК молекула, ДНК полимеразата се движи по веригата на майката, завършвайки изграждането на дъщерната верига според принципа на комплементарност.
Една от най-важните функции на ДНК полимеразата е да коригира грешките в новосинтезираната ДНК верига. Разпознавайки грешните базови двойки, ДНК полимеразата се връща една стъпка назад, грешната база се отстранява от дъщерната верига и се вмъква правилната. В резултат на това броят на грешките в репликацията на ДНК е приблизително една грешка на 10-100 милиарда базови двойки. Въпреки това, тъй като клетките се делят достатъчно често, грешките неизбежно се натрупват. Много от тези грешки засягат жизнеспособността на клетките. Ако клетките, в които е настъпила мутация, все пак оцелеят, имунната система ги унищожава с помощта на специални клетки, пригодени за тази цел.
Нека сега се върна към процеса на клетъчно делене. В профазата на митозата хромозомите се състоят от две хроматиди, което е резултат от репликация на ДНК в интерфазата. Не е възможно обаче да се регистрират под микроскоп, тъй като в профаза сестринските хроматиди са в близък контакт една с друга, като взаимно се спират една спрямо друга. В прометафазата ядрената мембрана се разрушава и хромозомите започват да се движат към екваториалната равнина на клетката. Този процес осигурява нормалното разделяне на хромозомите до полюсите на клетката по време на клетъчното делене, в резултат на което във всяка от новообразуваните клетки попада пълен набор от хромозоми. Сестринските хроматиди на една хромозома са свързани една с друга в точка, наречена центромер. В областта на центромера има кинетохор, сложна протеинова структура, свързана с ДНК на центромерната област. Филаментите на вретеното на деленето са прикрепени към тази зона по време на митоза. Както вече забелязахте, центриолите са разположени на полюсите на клетката, чийто брой също се удвоява преди клетъчното делене. Центриолите участват в образуването на филаментите на вретеното на деленето. Всяка нишка на вретено се прикрепя към една хромозома в областта на кинетохора. Във втората фаза на митозата, метафазата, хромозомите, състоящи се от две хроматиди, се подреждат в екваториалната равнина на клетката.
Филаментите на вретеното на деленето издърпват хромозоми, наречени от този момент дъщерни хромозоми, към противоположните краища на клетката. Фазата, през която хромозомите се движат в различни посоки, се нарича анафаза. Ако всичко върви по план, центриола се разделя и сестринските хроматиди се разминават към полюсите на клетката. Ако хромозомите не се подредят в екваториалната равнина на клетката в правилния ред, процесът на клетъчно делене може да бъде нарушен.
След като хромозомите се разпространят до противоположните краища на родителската клетка, клетката се разделя на две в резултат на цитокинеза. Четвъртият и последен етап на митозата се нарича телофаза. Клетките, които се размножават чрез митоза, както вече споменахме, се наричат соматични. Те включват всички клетки в нашето тяло, с изключение на клетките, които по-късно се превръщат в сперматозоиди или яйцеклетки. Клетките, които допринасят за образуването на нов организъм, се наричат зародишни клетки. Половите клетки се образуват в резултат на мейоза, която ще разгледаме в следващия раздел.
Митозата обикновено преминава без забавяне. Във всеки един момент обаче в човешкото тяло се извършват милиони клетъчни деления. От време на време в клетките има твърде много генетичен материал, докато в други може да е по-малко от нормалното. Обикновено тези клетки не оцеляват или се отстраняват от имунната система.
№85 Наследствени заболявания. Класификация на наследствените заболявания.Наследствени заболявания - заболявания, чието възникване и развитие е свързано с дефекти в софтуера на клетките, унаследени чрез гаметите. Терминът се използва по отношение на полиетиологични заболявания, за разлика от по-тясна група - генетични заболявания.
В зависимост от вида на първично засегнатите клетки се разграничават следните групи заболявания: заболявания, дължащи се на мутации в зародишните клетки – „гаметични“, т.е. всъщност наследствени заболявания (например фенилкетонурия, хемофилия); тези заболявания са наследствени; заболявания, дължащи се на мутации в соматичните клетки - "соматични" (например тумори, някои заболявания на имунна автоагресия). Тези заболявания не са наследствени; заболявания, дължащи се на комбинация от мутации в зародишни и соматични клетки (например семеен ретинобластом).
Леталните, сублеталните и хипогениталните заболявания се разглеждат отделно: смъртоносните заболявания водят до смърт по време на вътрематочното развитие (например монозомия по автозоми, хаплоидия, повечето полиплоидия); сублетални заболявания водят до смърт на индивида преди пубертета (например наследствени имунодефицити като швейцарски тип агам-магглобулинемия, синдром на Луи-Бар, някои хемофилии); хипогениталните заболявания се комбинират с безплодие (например синдроми на Шерешевски-Търнър, Клайнфелтер).
В зависимост от ролята на наследствеността и околната среда се разграничават четири групи заболявания: собствено наследствени заболявания (развиват се в резултат на мутации); екогенетични заболявания (развиват се при наличие на мутация под въздействието на специфична факторна среда); заболявания на генетично предразположение (приносът на генетичните фактори е очевиден); заболявания, причинени от фактори на околната среда (строго погледнато, тази група заболявания не принадлежи към наследствени заболявания, тъй като приносът на генетичните фактори за тяхното възникване, развитие и изход може да бъде изключително малък, съмнителен или неизвестен).