Mendel estava empenhado na seleção de ervilhas, e é às ervilhas, à sorte científica e ao rigor dos experimentos de Mendel que devemos a descoberta das leis básicas da herança: a lei da uniformidade dos híbridos de primeira geração, a lei da divisão e a lei da combinação independente.
Mendel fez experiências com 22 variedades de ervilhas, diferindo umas das outras em 7 características (cor, textura da semente). Mendel liderou seu trabalho por oito anos, estudou 10.000 pés de ervilha. Todas as formas de ervilhas que investigou eram de linhagem pura; os resultados do cruzamento dessas plantas entre si sempre foram os mesmos. Mendel apresentou os resultados de seu trabalho em um artigo de 1865, que se tornou a pedra angular da genética. É difícil dizer o que merece mais admiração nele e em seu trabalho - o rigor dos experimentos, a clareza da apresentação dos resultados, o perfeito conhecimento do material experimental ou o conhecimento do trabalho de seus predecessores.
Os colegas e contemporâneos de Mendel foram incapazes de avaliar a importância de suas conclusões. De acordo com A.E. Gaisinovich, até o final do século XIX. foi citado apenas cinco vezes e apenas um cientista - o botânico russo I.O. Schmalhausen - apreciou a importância deste trabalho. No entanto, no início do século 20, as leis por ele descobertas foram redescobertas quase simultaneamente e independentemente umas das outras pelos cientistas K. Correns, E. Cermak e K. de Vries. A importância dessas descobertas tornou-se imediatamente aparente para a comunidade científica no início do século XX; seu reconhecimento estava associado a certos sucessos em citologia e à formação da hipótese de hereditariedade nuclear.
|
Os principais padrões de herança de características. A hereditariedade humana tem sido estudada em uma extensão muito menor do que os mecanismos de herança de características em organismos vegetais e animais. No entanto, dados interessantes já foram obtidos sobre a dependência hereditária de muitas características físicas de uma pessoa. O nível moderno da ciência genética também nos permite afirmar que todos os padrões básicos de herança de traços e as leis da hereditariedade, identificados em experimentos com plantas e animais, também são válidos para humanos. Por exemplo, se um jovem que tem genes para olhos castanhos (homozigoto para castanhos) em ambos os cromossomos homólogos se casar com uma menina de olhos azuis cujos cromossomos também contêm apenas genes para olhos azuis, então seus filhos herdarão a cor dos olhos de acordo com as leis descobertas pelo fundador da genética G. Mendel nos anos 60 do século passado em seus experimentos com híbridos de plantas de ervilhas. A herança sexual em humanos e na maioria dos animais também ocorre de acordo com as leis gerais e está associada à distribuição de cromossomos que têm a forma de X e K (cromossomos sexuais). O conjunto de cromossomos do corpo feminino contém dois cromossomos X, e o corpo masculino contém um cromossomo X e um cromossomo Y (ver Fig. 7). O número de tais exemplos pode ser infinitamente grande, e não há dúvida de que o “retrato do gene” de uma pessoa (genótipo) determina em grande parte muitas de suas propriedades externas (fenótipo). Listadas abaixo estão algumas das características hereditárias de uma pessoa, determinadas por genes dominantes e recessivos (de acordo com K-Willie, 1974). A manifestação da influência do gene pode ser realizada em vários estágios da ontogênese, mas a maioria das características fenotípicas são determinadas antes mesmo do nascimento. Finalmente, a própria manifestação dos genes não é fatal, mas depende de fatores ambientais. Por exemplo, uma doença hereditária grave, fenilcetonúria, não se desenvolve se uma criança que contém os genótipos que causam essa doença no genótipo começa a receber uma determinada dieta imediatamente após o nascimento. Assim, a genética tem fatos convincentes que comprovam a existência de características estritamente hereditárias (por exemplo, níveis sanguíneos, hemofilia, cor do cabelo e dos olhos, características faciais e muitos outros) e características que são amplamente determinadas pelo ambiente externo (por exemplo, o crescimento e a massa de uma pessoa, a força e destreza de seus músculos, tendência a doenças, etc.). Deve-se notar que não há relação direta entre genes e características: o desenvolvimento de uma característica pode depender da influência de muitos genes, e um gene pode influenciar o desenvolvimento de muitas características. Esta imagem é ainda mais complicada pela correção constante do ambiente externo. Consequentemente, a propriedade da população e o desenvolvimento estão sempre intimamente relacionados. É impossível imaginar o desenvolvimento de uma criança sem as influências corretivas do meio ambiente, assim como é impossível sem um programa de desenvolvimento geneticamente predeterminado, que é uma síntese da experiência histórica de todas as gerações anteriores. Consideremos o papel da hereditariedade e do meio ambiente no desenvolvimento de algumas características físicas de uma pessoa no processo de seu desenvolvimento pré-natal e pós-natal. Desenvolvimento pré-natal. A formação dos órgãos e sistemas funcionais da criança em processo de embriogênese está sob o controle do genótipo, mas os fatores ambientais desempenham um papel importante. Para o embrião, o organismo materno é o ambiente primário. Aqui, em um órgão especial - o útero - o embrião está relativamente bem protegido de muitas influências nocivas e recebe tudo de que precisa para sua existência através da placenta. No entanto, especialmente nos estágios iniciais de desenvolvimento, muitos fatores que afetam o corpo materno também afetam o desenvolvimento do embrião (os mais significativos são: radiação ionizante, doenças sofridas pela mulher durante a gravidez e muitos produtos químicos: álcool, nicotina, antibióticos, drogas hormonais, etc.). Deve-se notar que no período pré-natal de uma pessoa existem períodos críticos em que o organismo em desenvolvimento é especialmente sensível à ação de fatores externos. Existem dois desses períodos. O primeiro inclui o início do desenvolvimento pré-natal, suas primeiras três semanas. Nesse momento, ocorre a postura de todos os órgãos mais importantes, e os efeitos adversos durante esse período geralmente levam à morte do embrião. Durante o segundo período crítico (da 4ª à 7ª semana), ocorre um maior desenvolvimento de todos os órgãos e influências externas prejudiciais durante este período podem levar ao nascimento de uma criança com vários defeitos físicos (Fig. 9). É importante que os mesmos defeitos físicos congênitos possam estar associados a danos às estruturas genéticas e à ação de fatores externos desfavoráveis no processo de embriogênese. Esta é uma boa evidência de que o meio ambiente e a hereditariedade são igualmente responsáveis pelo desenvolvimento normal da criança. Portanto, no momento do nascimento, todos os órgãos de uma pessoa e seus sistemas fisiológicos, incluindo o sistema nervoso, estão geralmente formados. Naturalmente, surge a pergunta: as propriedades do sistema nervoso que subjazem à atividade mental de uma pessoa, a base de sua mente, são predeterminadas hereditariamente? Existem "preparações morfológicas" prontas para a consciência, da mesma maneira que existem para a atividade nervosa inferior associada à regulação dos processos vitais de nosso corpo? Para responder a essas perguntas, vamos nos deter no próximo estágio pós-natal do desenvolvimento humano. Desenvolvimento infantil pós-natal. Em primeiro lugar, consideraremos os principais métodos que nos permitem estudar a importância específica do meio ambiente e da hereditariedade no desenvolvimento pós-natal de crianças e adolescentes. Realizar tais experimentos em plantas é simples e consiste em isolar dois grupos de organismos, idênticos em genótipo, e colocá-los em diferentes condições de existência. Por exemplo, um grupo de plantas (controle) é cultivado em condições normais, outro (experimental) - em uma sala escura. Como resultado de tais experimentos, pode-se concluir que a formação de clorofila verde nas plantas depende não apenas da hereditariedade, mas também de fatores ambientais (luz), uma vez que plantas cultivadas no escuro não conterão esse pigmento. Realizar experimentos semelhantes em humanos é impossível tanto do ponto de vista moral e ético quanto do biológico. Não existem duas pessoas no mundo com os mesmos genótipos. Mas ... há exceções à regra. Estes são gêmeos idênticos ou homozigotos (IB), tendo não apenas semelhanças externas impressionantes, mas também "retratos" de genes quase idênticos. O nascimento de gêmeos idênticos e não idênticos (TW) não é incomum; em média, uma em cada 100 gestações termina no nascimento de mais de um filho. Gêmeos é o "material" mais excelente apresentado ao cientista pela própria natureza, portanto o método em si é chamado de gêmeo. Qual é a sua essência? Digamos que você precise estabelecer qual fator é o mais responsável pelo desenvolvimento físico e crescimento de uma criança: hereditariedade ou meio ambiente? Para tanto, são identificados casais gêmeos, realizado seu exame morfológico e estudado seu estilo de vida (geralmente gêmeos, principalmente idênticos, têm interesses semelhantes e condições de criação próximas). Com base nos estudos realizados, estabelece-se o coeficiente de semelhança (concordância) entre gêmeos e conclui-se sobre o papel da hereditariedade ou do ambiente no desenvolvimento de certas qualidades. Os resultados de tais estudos de gêmeos idênticos e não idênticos em termos da frequência de ocorrência de algumas doenças entre eles são apresentados na tabela. Infelizmente, os professores ainda estão fazendo pouco trabalho para identificar crianças superdotadas, o que muitas vezes está associado a uma subestimação da individualidade fisiológica e mental das crianças, com o desconhecimento da biologia elementar de uma criança. O fraco trabalho na identificação da superdotação infantil também pode ser explicado pelo ainda baixo nível de pesquisas nessa área e pela falta de critérios científicos precisos pelos quais seria fácil para um professor detectar as inclinações hereditárias de uma criança. Existe apenas uma conexão estreita entre superdotação e um alto nível de excitabilidade do sistema nervoso, um sintoma do qual muitas vezes é um desequilíbrio acentuado (psicopaticidade) da criança. O professor deve estar especialmente atento a essas crianças. Isso é necessário principalmente do lado higiênico, uma vez que as ações erradas do professor podem levar ao desenvolvimento de traços de caráter extremos em patológicos. Também é necessário para a identificação oportuna de inclinações hereditárias e seu desenvolvimento ideal. É importante lembrar que a aparente falta de vontade de aprender, a preguiça exterior e a indisciplina não são evidências suficientes de uma falta de habilidade. Há muitas evidências na história de que essa conclusão é verdadeira. O criador da teoria da relatividade A. Einstein na infância não mostrou habilidades abertas e somente aos 9 anos foi capaz de ir para o ensino preparatório. A. Humboldt (1769-1859) e L. Pasteur (1822-1895), o notável físico inglês I. Newton, eram conhecidos como alunos preguiçosos e incapazes na escola. Pode-se supor que o tratamento habilidoso deles por pais e professores criou, no final, todas as condições necessárias para o desenvolvimento de talentos. Além disso, a qualidade mais importante do gênio é o trabalho árduo. O grande inventor americano T. Edison escreveu: "O gênio é um por cento da inspiração e noventa e nove por cento está suando." Quem não aprende a "suar" nunca poderá obter grande êxito em qualquer tipo de atividade humana, apesar das inclinações hereditárias mais marcantes. Quem, senão um professor, deveria antes de tudo educar a laboriosidade na criança - este é o fator mais importante no desenvolvimento progressivo da personalidade, a principal condição para a formação de quaisquer habilidades. Assim, a hereditariedade fornece apenas "matéria-prima", e o ambiente realiza seu principal "processamento". Uma pessoa que nasceu mesmo com as inclinações mais favoráveis, mas vive em um ambiente que impede o desenvolvimento de suas habilidades, continuará a ser uma mediocridade, de modo que a responsabilidade principal pelo desenvolvimento das habilidades intelectuais da criança recai sobre os educadores. No entanto, o papel do meio ambiente não deve ser absolutizado, não se deve esquecer que o “material” fornecido pela hereditariedade nem sempre é de alta qualidade, e mesmo o escultor mais talentoso não será capaz de esculpir uma escultura em areia, apenas já que um bloco de mármore por si só não se tornará uma obra de arte ... №67 - nº 68. Herança autossômica dominante- um tipo de herança em que um alelo mutante localizado no autossomo é suficiente para a doença (ou traço) se expressar. Herança autossômica recessiva- o tipo de herança de um traço ou doença, em que um alelo mutante localizado em um autossomo deve ser herdado de ambos os pais. |
№69. Herança ligada ao sexo - herança de um gene localizado nos cromossomos sexuais. A herança de características que aparecem apenas em indivíduos do mesmo sexo, mas não são determinadas por genes localizados nos cromossomos sexuais, é chamada de herança limitada por sexo.
A herança ligada ao cromossomo X é a herança de genes quando o sexo masculino é heterogamético e se caracteriza pela presença de um cromossomo Y (XY), e as mulheres são homogaméticas e possuem dois cromossomos X (XX). Todos os mamíferos (incluindo humanos), a maioria dos insetos e répteis possuem esse tipo de herança.
№70 Tipo de herança holandesa. Herança holandric (herança holandric) - herança de uma característica controlada por um gene localizado em uma parte não homóloga do cromossomo Y. O holandricgene é um gene ligado ao cromossomo Y masculino. Os sinais determinados por G. g são transmitidos apenas pela linha masculina - de pai para filho.
№71 Com tipo de herança ligada ao X o gene mutante está localizado no cromossomo X. Se a mutação tiver um efeito dominante, tanto homens quanto mulheres podem ficar doentes. No entanto, a doença é transmitida de um pai doente com probabilidade de 100% apenas para meninas, mas não para meninos, que recebem um cromossomo Y de seu pai (Fig. 5). A probabilidade de transmissão de uma mutação dominante ligada ao X de uma mãe doente para os filhos é de 50%. A doença tem a mesma probabilidade de ser herdada tanto pela filha quanto pelo filho.
Para o dominante,ligado ao cromossomo X, o tipo de herança é a patologia conhecida pelos pediatras, raquitismo resistente à vitamina D (sinônimos: hipofosfatemia, hipofosfatemia familiar ligada ao X, diabetes fosfático). O diagnóstico desse raquitismo severo, que não desaparece sob a influência de grandes doses de vitamina D, é confirmado pela presença de doença semelhante em alguns parentes, homens e mulheres.
Mais frequentemente doenças ligadas ao Xsão herdados de forma recessiva. Uma característica distintiva das doenças com recessivaTipo de herança ligada ao X é que os homens estão doentes na família e herdam o alelo mutante de sua mãe praticamente saudável, heterozigota para o alelo mutante. Ao compilar um gráfico de linhagem, essas mães costumam ter irmãos ou tios doentes. Homens doentes podem transmitir sua doença apenas por uma geração e apenas para netos (mas não netas) por meio de sua filha saudável, mas heterozigota.
Assim, se você traçar na herança do pedigree na linha masculinadoença recessiva ligada ao X, você obtém algo como um "movimento do cavaleiro de xadrez".
Maioriadoenças conhecidas ligadas ao Xestãohemofilia A e B , bem como a patologia mais grave do sistema muscular -Distrofia muscular de Duchenne Becker ... O desenvolvimento da hemofilia A é baseado em mutações no gene responsável pela síntese do fator VIII da coagulação do sangue, e na hemofilia B, o fator IX da coagulação do sangue é defeituoso. Ambos os genes que codificam os fatores VIII e IX estão localizados no braço longo do cromossomo X em q28 e q27.1-2, respectivamente. Sabe-se que na hemofilia há violação da coagulação sanguínea, e os menores cortes podem levar à morte um paciente sem cuidados hematológicos especiais. Observe que nas mulheres - portadoras do gene da hemofilia (os chamados "condutores"), em alguns casos, há também tendência ao sangramento, que se expressa em menstruações abundantes e sangramento prolongado durante o parto. Essa circunstância deve ser levada em consideração pelos ginecologistas-obstetras ao trabalharem com mulheres portadoras de alelos mutantes em qualquer um dos genes A ou B da hemofilia.
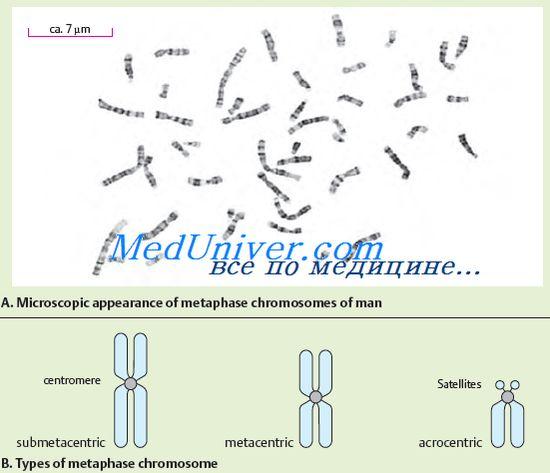
№72 A estrutura e função dos cromossomos. A morfologia cromossômica é melhor observada na célula no estágio de metáfase. O cromossomo consiste em dois corpos em forma de bastão - cromátides. Ambas as cromátides de cada cromossomo são geneticamente idênticas entre si. Os cromossomos são diferenciados em comprimento. Os cromossomos têm um centrômero ou constrição primária, dois telômeros e dois braços. Em alguns cromossomos, constrições secundárias e satélites são isolados. O movimento do cromossomo determina o centrômero, que possui uma estrutura complexa. O centrômero de DNA se distingue por sua sequência de nucleotídeos característica e proteínas específicas. Dependendo da localização do centrômero, cromossomos acrocêntricos, submetacêntricos e metacêntricos são diferenciados. Como mencionado acima, alguns cromossomos têm constrições secundárias. Eles, ao contrário da constrição primária (centrômero), não servem como local de fixação dos filamentos do fuso e não desempenham nenhum papel no movimento dos cromossomos. Algumas constrições secundárias estão associadas à formação de nucléolos, neste caso são chamados de organizadores nucléolos. Os organizadores nucleolares contêm os genes responsáveis pela síntese de RNA. A função de outras constrições secundárias ainda não está clara. Alguns cromossomos acrocêntricos possuem satélites - regiões que são conectadas ao resto do cromossomo por um fino fio de cromatina. A forma e o tamanho do satélite são constantes para um determinado cromossomo. Em humanos, existem satélites em cinco pares de cromossomos. As extremidades dos cromossomos ricos em heterocromatina estrutural são chamadas de telômeros. Os telômeros evitam que as extremidades dos cromossomos se colem após a reduplicação e, assim, contribuem para a preservação de sua integridade. Conseqüentemente, os telômeros são responsáveis pela existência de cromossomos como entidades individuais. Os cromossomos que possuem a mesma ordem de genes são chamados de homólogos. Eles têm a mesma estrutura (comprimento, localização do centrômero, etc.). Os cromossomos não homólogos têm um conjunto de genes diferente e uma estrutura diferente. O exame da estrutura fina dos cromossomos mostrou que eles são compostos de DNA, proteína e uma pequena quantidade de RNA. A molécula de DNA carrega cargas negativas distribuídas ao longo de todo o seu comprimento, e as proteínas ligadas a ela - as histonas são carregadas positivamente. Este complexo DNA-proteína é denominado cromatina. A cromatina pode ter vários graus de condensação. A cromatina condensada é chamada de heterocromatina, a cromatina descondensada é chamada de eucromatina. O grau de descondensação da cromatina reflete seu estado funcional. As regiões de heterocromatina são funcionalmente menos ativas do que as regiões de eucromatina, nas quais a maioria dos genes está localizada. Faça a distinção entre heterocromatina estrutural, cuja quantidade difere em diferentes cromossomos, mas está constantemente localizada nas regiões quase centroméricas. Além da heterocromatina estrutural, há uma heterocromatina opcional que aparece no cromossomo durante o superenrolamento de regiões eucromáticas. A confirmação da existência desse fenômeno nos cromossomos humanos é o fato da inativação genética de um cromossomo X nas células somáticas de uma mulher. A sua essência reside no facto de existir um mecanismo de inactivação evolutivamente formado da segunda dose de genes localizados no cromossoma X, pelo que, apesar do número diferente de cromossomas X nos organismos masculinos e femininos, o número de genes funcionar neles é igual. A cromatina é condensada ao máximo durante a divisão celular mitótica, então pode ser detectada na forma de cromossomos densos.As dimensões das moléculas de DNA dos cromossomos são enormes. Cada cromossomo é representado por uma molécula de DNA. Eles podem atingir centenas de micrômetros e até centímetros. Dos cromossomos humanos, o maior é o primeiro; seu DNA tem comprimento total de até 7 cm. O comprimento total das moléculas de DNA de todos os cromossomos de uma célula humana é de 170 cm. Apesar do tamanho gigantesco das moléculas de DNA, ele é bastante denso em cromossomos. Este empacotamento específico de DNA cromossômico é fornecido por proteínas histonas. As histonas são organizadas ao longo do comprimento da molécula de DNA na forma de blocos. Um bloco contém 8 moléculas de histonas, formando um nucleossomo (uma formação que consiste em uma fita de DNA enrolada em torno de um octâmero de histona). O nucleossomo tem cerca de 10 nm de tamanho. Os nucleossomos parecem contas amarradas em um cordão. Os nucleossomos e as seções de DNA que os conectam são compactados na forma de uma espiral; há seis nucleossomos para cada volta dessa espiral. É assim que a estrutura do cromossomo é formada. A informação hereditária de um organismo é estritamente ordenada de acordo com os cromossomos individuais. Cada organismo é caracterizado por um conjunto específico de cromossomos (número, tamanho e estrutura), que é denominado cariótipo. O cariótipo humano é representado por vinte e quatro cromossomos diferentes (22 pares de autossomos, cromossomos X e Y). O cariótipo é o passaporte da espécie. A análise do cariótipo pode detectar distúrbios que podem levar a anormalidades do desenvolvimento, doenças hereditárias ou morte de fetos e embriões nos estágios iniciais de desenvolvimento. Por muito tempo, acreditou-se que o cariótipo humano consistia em 48 cromossomos. Porém, no início de 1956, foi publicado um relatório segundo o qual o número de cromossomos no cariótipo humano é de 46. Os cromossomos humanos diferem em tamanho, localização dos centrômeros e constrições secundárias. Pela primeira vez, a divisão do cariótipo em grupos foi realizada em 1960 em um congresso em Denver (EUA). A descrição do cariótipo humano originalmente incluía os seguintes dois princípios: a localização dos cromossomos ao longo de seu comprimento; agrupamento de cromossomos por localização do centrômero (metacêntrico, submetacêntrico, acrocêntrico). A constância exata do número de cromossomos, sua individualidade e a complexidade da estrutura indicam a importância da função que desempenham. Os cromossomos desempenham a função do principal aparato genético da célula. Neles, os genes são organizados em uma ordem linear, cada um dos quais ocupa um lugar estritamente definido (locus) no cromossomo. Cada cromossomo contém muitos genes, mas para o desenvolvimento normal de um organismo, um conjunto de genes de um conjunto completo de cromossomos é necessário.
№73. A hereditariedade se baseia na capacidade de todos os organismos vivos de acumular, armazenar e transmitir informações hereditárias aos descendentes. Esta uma das características qualitativas mais importantes da matéria viva está associada aos ácidos nucléicos - desoxirribonucléico (DNA) e ribonucléico (RNA). O papel principal pertence ao DNA - a molécula mais longa dos organismos vivos, concentrada nos núcleos das células e representando seu aparato hereditário. O grande comprimento da molécula de DNA permite "escrever" nela, como em uma fita telegráfica, todas as propriedades básicas do futuro organismo e o programa de seu desenvolvimento. Tal "registro" é realizado usando uma "linguagem nucleica" especial, ou "código nucleico", cuja essência é alterar a sequência dos quatro compostos químicos que constituem o DNA. Figurativamente falando, a "linguagem nucleica" consiste de quatro letras a partir das quais palavras separadas são construídas e frases inteiras da "linguagem nucléica".
Em tal "faixa" nucleica, áreas independentes separadas podem ser distinguidas, incluindo uma descrição do programa de desenvolvimento de uma característica. Eles são chamados de genes. Cada molécula de DNA inclui centenas de genes e é um programa para o desenvolvimento de muitas características e propriedades do organismo. Combinando-se com proteínas especiais e algumas outras substâncias, as moléculas de DNA formam formações especiais no núcleo - os chamados cromossomos.
Organização do material genético. Genes [do grego. genos, nascimento] - uma unidade de hereditariedade, um pedaço de DNA que ocupa um lugar específico no cromossomo. Do ponto de vista da genética, um gene é um fator herdado e uma unidade indivisível de material genético. O gene estrutural (cistron) é um fragmento de DNA envolvido na formação de uma cadeia polipeptídica. Consiste em uma sequência líder, fragmentos codificadores (exões), sequências de inserção (nitrões) e uma sequência terminal. Uma vez que algumas proteínas consistem em mais de uma subunidade, a expressão "um gene - uma enzima" em relação a uma proteína heteromultimérica (isto é, consistindo em duas ou mais subunidades polipeptídicas diferentes) deve ser interpretada como "um gene - uma cadeia polipeptídica" .
O genótipo é uma coleção de genes em um organismo. Mesmo nos tempos antigos, as pessoas usavam empiricamente as leis da herança. Com base nessa experiência, a seleção foi desenvolvida [de lat. selectio, choose] - a ciência dos métodos de criação de novas variedades de plantas e raças de animais por meio de seleção e cruzamento. Até recentemente, o genótipo parecia inexpugnável, não sujeito às ações humanas. A descoberta da estrutura dos genes possibilitou isolá-los de forma isolada, sintetizá-los bioquimicamente e até introduzi-los no corpo. Tornou-se possível influenciar o gene sem excretá-lo do corpo. Tudo isso criou os pré-requisitos para a manipulação do genótipo.
№74 A estrutura dos ácidos nucléicos.
Ácido nucleico(do latim núcleo - núcleo) - um composto orgânico de alto peso molecular, um biopolímero (polinucleotídeo) formado por resíduos de nucleotídeos. Os ácidos nucléicos DNA e RNA estão presentes nas células de todos os organismos vivos e desempenham as funções mais importantes de armazenamento, transmissão e implementação de informações hereditárias. As formas poliméricas de ácidos nucléicos são chamadas de polinucleotídeos. As cadeias de nucleotídeos são conectadas por meio de um resíduo de ácido fosfórico (ligação fosfodiéster). Uma vez que existem apenas dois tipos de moléculas heterocíclicas nos nucleotídeos, ribose e desoxirribose, existem apenas dois tipos de ácidos nucléicos - ácido desoxirribonucléico (DNA) e ácido ribonucléico (RNA).
As formas monoméricas também são encontradas nas células e desempenham um papel importante na sinalização ou no armazenamento de energia. O monômero de RNA mais famoso é o ATP, ácido adenosina trifosfórico, o mais importante acumulador de energia na célula.
Uma das principais funções dos ácidos nucléicos é determinar a síntese de proteínas. As informações sobre a estrutura das proteínas codificadas na sequência de nucleotídeos do DNA devem ser transmitidas de uma geração para outra e, portanto, devem ser copiadas corretamente, ou seja, síntese exatamente da mesma molécula de DNA (replicação).
№75 RepetirLigação de DNA o processo de síntese de uma molécula filha de ácido desoxirribonucléico na matriz da molécula de DNA original. Durante a divisão subsequente da célula-mãe, cada célula-filha recebe uma cópia da molécula de DNA, que é idêntica ao DNA da célula-mãe original. Este processo garante a transmissão precisa da informação genética de geração em geração. A replicação do DNA é realizada por um complexo enzimático complexo que consiste em 15-20 proteínas diferentes, denominado repissoma. A replicação do DNA é um evento chave na divisão celular. É essencial que no momento da divisão o DNA seja totalmente replicado e apenas uma vez. Isso é fornecido por certos mecanismos de regulação da replicação do DNA. A replicação ocorre em três estágios:
iniciação de replicação
alongamento
término da replicação.
A regulação da replicação é realizada principalmente no estágio de iniciação. Isso é muito fácil de implementar, porque a replicação pode começar não a partir de qualquer pedaço de DNA, mas de um DNA estritamente definido, denominado local de iniciação da replicação. Pode haver apenas um ou vários desses sites no genoma. O conceito de replicon está intimamente relacionado ao conceito de um local de início de replicação. Um replicon é um trecho de DNA que contém um local de iniciação da replicação e se replica assim que a síntese de DNA começa nesse local. Os genomas das bactérias, via de regra, representam um replicon, o que significa que a replicação de todo o genoma é o resultado de apenas um ato de iniciação da replicação. Os genomas dos eucariotos (assim como seus cromossomos individuais) consistem em um grande número de replicons independentes, o que reduz significativamente o tempo total de replicação de um cromossomo individual. Os mecanismos moleculares que controlam o número de atos de iniciação da replicação em cada local durante um ciclo de divisão celular são chamados de controle do número de cópias. Além do DNA cromossômico, as células bacterianas geralmente contêm plasmídeos, que são replicons individuais. Os plasmídeos têm seus próprios mecanismos de controle de cópia: eles podem fornecer a síntese de apenas uma cópia do plasmídeo por ciclo celular, ou milhares de cópias.
A replicação começa no local de início da replicação com o desenrolamento da dupla hélice do DNA, com a formação de uma forquilha de replicação - o local da replicação direta do DNA. Cada site pode formar um ou dois bifurcações de replicação, dependendo se a replicação é unidirecional ou bidirecional. A replicação bidirecional é mais comum. Algum tempo após o início da replicação em um microscópio eletrônico, é possível observar o olho da replicação - uma seção do cromossomo onde o DNA já foi replicado, circundada por seções mais extensas de DNA não replicado.
No garfo de replicação, o DNA copia um grande complexo de proteínas (replicissoma), cuja enzima chave é a DNA polimerase. A bifurcação de replicação se move a uma taxa de cerca de 100.000 pares de bases por minuto em procariotos e 500-5.000 em eucariotos.
№76 A estrutura e tipos de cromossomos.O núcleo de cada célula somática do corpo humano contém46 cromossomos... Kitcromossomos cada indivíduo, normal e patológico, é denominado cariótipo.
A partir de46 cromossomosque compõem o conjunto de cromossomos humanos, 44 ou 22 pares representam autossômicoscromossomos , o último par são cromossomos sexuais. Nas mulheres, a constituição dos cromossomos sexuais é normalmente representada por dois cromossomos X, e nos homens - pelos cromossomos X e Y.
Em tudopares de cromossomos tanto autossômico quanto sexual, um dos cromossomos é obtido do pai e o outro da mãe. Os cromossomos de um par são chamados de homólogos ou cromossomos homólogos. As células reprodutivas (espermatozoides e óvulos) contêm um conjunto haplóide de cromossomos, ou seja, 23 cromossomos.
 Os espermatozoides são divididos em dois tipos, dependendo se contêmcromossomaX ou Y. Todos os ovos normalmente contêmapenas cromossomo
X.
Os espermatozoides são divididos em dois tipos, dependendo se contêmcromossomaX ou Y. Todos os ovos normalmente contêmapenas cromossomo
X.
Cromossomos são claramente visíveis após coloração especial durante a divisão celular, quando os cromossomos são espiralizados ao máximo. Nesse caso, uma constrição é detectada em cada cromossomo, que é chamada de centrômero. O centrômero divide o cromossomo em um braço curto (denotado pela letra "p") e um braço longo (denotado pela letra "q"). O centrômero determina o movimento do cromossomo durante a divisão celular. De acordo com a posição do centrômero, os cromossomos são classificados em vários grupos. Se o centrômero estiver localizado no meio do cromossomo, esse cromossomo é denominado metacêntrico; se o centrômero estiver localizado próximo a uma das extremidades do cromossomo, será denominado acrocêntrico. Alguns cromossomos acrocêntricos têm os chamados satélites, que formam nucléolos em uma célula que não se divide. Os nucléolos contêm numerosas cópias de rRNA. Além disso, os cromossomos submetacêntricos são diferenciados, quando o centrômero não está localizado no meio do cromossomo, mas um tanto deslocado para uma das extremidades, mas não tão significativamente quanto nos cromossomos acrocêntricos.
As pontas de cadacromossomo do ombro chamados telômeros. Verificou-se que os telômeros desempenham um papel importante na manutenção da estabilidade dos cromossomos. Os telômeros contêm um grande número de repetições da sequência de nucleotídeos TTAGGG, as chamadas repetições em tandem. Normalmente, durante a divisão celular, há uma diminuição no número dessas repetições em telômeros .
No entanto, cada vez que são concluídos com a ajuda de uma enzima especial chamadatelomerase... Uma diminuição na atividade dessa enzima leva ao encurtamento dos telômeros, que se acredita ser a causa da morte celular e normalmente acompanha o envelhecimento.
№77. Classificação dos cromossomos de Denver. A classificação e a nomenclatura de cromossomos humanos uniformemente corados foram adotadas pela primeira vez em um encontro internacional em 1960 em Denver, mais tarde modificadas e complementadas (Londres, 1963 e Chicago, 1966). De acordo com a classificação de Denver, todos os cromossomos humanos são divididos em 7 grupos, dispostos em ordem decrescente de seu comprimento e levando em consideração o índice do centríolo (a razão entre o comprimento do braço curto e o comprimento de todo o cromossomo, expresso como um percentagem). Os grupos são designados por letras do alfabeto inglês de A a G. Todos os pares de cromossomos são geralmente numerados com algarismos arábicos. As características dos grupos são apresentadas na tabela. 4. A classificação proposta tornou possível distinguir claramente entre cromossomos pertencentes a grupos diferentes. Desde 1960, o rápido desenvolvimento da citogenética clínica começa: em 1959, J. Lejeune descobriu a natureza cromossômica da síndrome de Down; K. Ford, P. Jacobs e J. Strong descreveram as características do cariótipo nas síndromes de Klinefelter e Turner; no início dos anos 70. a natureza cromossômica das síndromes de Edwards e Patau, a síndrome do "grito do gato" foi descoberta; descreveu instabilidade cromossômica em várias síndromes hereditárias e doenças malignas. Ao mesmo tempo, a aplicação do método de obtenção de cromossomos uniformemente coloridos revelou-se insuficientemente eficaz para a identificação de cromossomos. A desvantagem da classificação de Denver é que a diferenciação de pares homólogos dentro de um grupo de cromossomos frequentemente encontra dificuldades intransponíveis.
Tabela 4^ Grupos de cromossomos e suas características
|
Grupo |
Número do cromossomo |
Localização do centrômero |
Índice centríolo (%) |
Observação |
|
O maior metacêntrico |
No ombro longo, pode haver uma constrição secundária |
|||
|
O maior submetacêntrico | ||||
|
Grande metacêntrico |
20% mais curto que o primeiro |
|||
|
Grande submetacêntrico | ||||
|
6-12 e cromossomo X |
Submetacêntrico médio |
No dia 9, costuma haver uma constrição secundária |
||
|
Acrocêntrico médio |
Em todas as constrições secundárias |
|||
|
Metacêntrico pequeno |
Constrição secundária ocorre em 10% dos casos. |
|||
|
Pequenos submetacêntricos | ||||
|
Menor metacêntrico | ||||
|
21-22 e cromossomo Y |
Menor acrocêntrico |
Nos dias 21 e 22 constrições secundárias |
№78 Biossíntese de proteínas na célula. A síntese de proteínas é um processo muito complexo de várias etapas. Ocorre em organelas especiais - ribossomos. A célula contém um grande número de ribossomos. Por exemplo, E. coli tem cerca de 20.000 deles.
Como ocorre a síntese de proteínas nos ribossomos?
As moléculas de proteína são essencialmente cadeias polipeptídicas compostas por aminoácidos individuais. Mas os aminoácidos não são ativos o suficiente para se ligarem por conta própria. Portanto, antes de se conectar e formar uma molécula de proteína, os aminoácidos devem ser ativados. Essa ativação ocorre sob a ação de enzimas especiais. Além disso, cada aminoácido tem sua própria enzima sintonizada especificamente para ele.
A fonte de energia para isso (como para muitos processos na célula) é trifosfato de adenosina (ATP).
Como resultado da ativação, o aminoácido torna-se mais lábil e, sob a ação da mesma enzima, liga-se ao t-RNA.
É importante que cada aminoácido corresponda a um t-RNA estritamente específico. Ela encontra "seu" aminoácido e o transfere para o ribossomo. Portanto, esse RNA foi denominado RNA de transporte.
Consequentemente, vários aminoácidos ativados, combinados com seus t-RNAs, entram no ribossomo. O ribossomo é, por assim dizer, uma correia transportadora para montar uma cadeia de proteínas a partir de vários aminoácidos que entram nela.
Acontece que simultaneamente com o t-RNA, no qual seu próprio aminoácido "se assenta", o ribossomo recebe um "sinal" do DNA contido no núcleo. De acordo com este sinal, uma ou outra proteína, uma ou outra enzima é sintetizada no ribossomo (visto que enzimas são proteínas).
A influência direta do DNA na síntese de proteínas não é realizada diretamente, mas com a ajuda de um mediador especial, aquela forma de RNA, que é chamada de RNA mensageiro ou mensageiro (m-RNA ou i-RNA).
O RNA mensageiro é sintetizado no núcleo pelo iodo pela influência do DNA, portanto sua composição reflete a composição do DNA. A molécula de RNA é como um molde em forma de DNA.
O m-RNA sintetizado entra no ribossomo e, por assim dizer, transmite a essa estrutura um plano - em que ordem os aminoácidos ativados que entram no ribossomo devem ser combinados entre si para que uma certa proteína seja sintetizada. Caso contrário, a informação genética codificada no DNA é transferida para o m-RNA e então para a proteína.
№79 Mutação (latmutação - mudança) - alteração persistente (isto é, que pode ser herdada pelos descendentes de uma determinada célula ou organismo) no genótipo que ocorre sob a influência do ambiente externo ou interno. Termo cunhado por Hugo de Vries. O processo de ocorrência de mutações é denominado mutagênese.
Genômica: - poliploidização (a formação de organismos ou células, cujo genoma é representado por mais de dois (3n, 4n, 6n, etc.) conjuntos de cromossomos) e neuploidia (heteroploidia) - uma mudança no número de cromossomos que não é um múltiplo do conjunto haplóide (ver Inge-Vechtomov, 1989). Dependendo da origem dos conjuntos de cromossomos, os poliplóides distinguem entre alopoliplóides, que possuem conjuntos de cromossomos obtidos por hibridização de diferentes espécies, e autopoliplóides, que apresentam um aumento no número de conjuntos de cromossomos de seu próprio genoma, um múltiplo de n.
Com as mutações cromossômicas, ocorrem grandes mudanças na estrutura dos cromossomos individuais. Nesse caso, ocorre a perda (deleção) ou duplicação de uma parte (duplicação) do material genético de um ou vários cromossomos, alteração na orientação dos segmentos cromossômicos em cromossomos individuais (inversão), bem como a transferência de uma parte do material genético de um cromossomo para outro (translocação) (um caso extremo - a união de cromossomos inteiros, a chamada translocação robertsoniana, que é uma opção de transição de mutação cromossômica para genômica).
No nível do gene, as mudanças na estrutura primária do DNA dos genes sob a influência de mutações são menos significativas do que com as mutações cromossômicas; no entanto, as mutações genéticas são mais comuns. Como resultado de mutações genéticas, substituições, deleções e inserções de um ou mais nucleotídeos, translocações, duplicações e inversões de várias partes do gene ocorrem. No caso em que apenas um nucleotídeo muda sob a ação de uma mutação, fala-se de mutações pontuais. Uma vez que o DNA contém apenas dois tipos de bases nitrogenadas - purinas e pirimidinas, todas as mutações pontuais com substituições de base são divididas em duas classes: transições (substituição de purina por purina ou pirimidina por pirimidina) e transversão (substituição de purina por pirimidina ou vice-versa) . Existem quatro consequências genéticas possíveis para as mutações pontuais: 1) preservação do significado do códon devido à degenerescência do código genético (terminação sinônima (mutação sem sentido). Existem três códons sem sentido no código genético: âmbar - UAG, ocp - UAA e opala - UGA (de acordo com este, o nome e as mutações que levam à formação de trigêmeos sem sentido - por exemplo, mutação âmbar), 4) reverso substituição (códon de parada com códon sentido).
De acordo com sua influência na expressão gênica, as mutações são divididas em duas categorias: mutações do tipo de substituições de pares de bases e do tipo de deslocamento de quadro (frameshift). Os últimos são deleções ou inserções de nucleotídeos, cujo número não é múltiplo de três, o que está associado à natureza tripla do código genético.
Uma mutação primária é às vezes chamada de mutação direta, e uma mutação que restaura a estrutura original de um gene é chamada de mutação reversa ou reversão. O retorno ao fenótipo original do organismo mutante devido à restauração da função do gene mutante muitas vezes ocorre não devido a uma reversão verdadeira, mas devido a uma mutação em outra parte do mesmo gene ou mesmo em outro gene não alélico. Nesse caso, a mutação recorrente é chamada de mutação supressora. Os mecanismos genéticos pelos quais o fenótipo mutante é suprimido são muito diversos.
Mutações renais (esporos) são mutações somáticas persistentes que ocorrem nas células dos pontos de crescimento das plantas. Leva a variação clonal. Eles são preservados durante a propagação vegetativa. Muitos cultivares são mutações renais.
№80. Empacotando DNA em um cromossomo. Os cromossomos são compostos de cromatina - uma mistura de DNA e proteínas em uma proporção de ~ 1: 1. É devido à interação com as proteínas que a molécula de DNA tem uma estrutura compacta. O comprimento médio de uma fita de DNA humano (se esticada) é de 4 cm, ou seja, o núcleo de cada célula contém cerca de 1 metro de DNA. O empacotamento de uma molécula de DNA em um cromossomo tem três níveis (veja a figura): 1. nucleossomos, uma fita do nucleossomo 2. hélice do solenóide 3. loops Uma dupla hélice de DNA conhecida por cada um de seus tipos "enrolada" em torno de um complexo de 4 pares de moléculas de proteína forma um "grânulo" - nucleossomo. Os nucleossomos, interligados por seções da molécula de DNA, formam um fio, que por sua vez é trançado em forma de solenóide (conforme um dos modelos existentes), de uma volta - 6 nucleossomos. Essa estrutura forma laços no próximo nível de embalagem. Nessa forma, o DNA está presente no núcleo celular na fase de crescimento celular, síntese de seus componentes, incluindo a duplicação do DNA. No entanto, em diferentes estágios da vida celular, a condensação da cromatina é diferente. Durante o período de crescimento celular, a cromatina no núcleo não se condensa, a compactação de sua localização difere para as diferentes partes do cromossomo. Quando ocorre o processo de divisão celular, a cromatina torna-se cada vez mais densa e as alças, mencionadas acima, formam "rosetas" compactadas uma na outra (também segundo um dos modelos existentes), 18 alças por anel. Como resultado, a cromatina assume a forma de cromossomos condensados, tão conhecidos por nós nos livros didáticos.
№81. Gametogênese.(do Gameta e da gênese grega - origem) o processo de desenvolvimento e formação das células sexuais - gametas (ver Gametas). G. gametas masculinos (espermatozóides, espermatozoides) são chamados de espermatogênese, gametas femininos (oócitos) - oogênese. Em animais e plantas, G. procede de forma diferente, dependendo do lugar da meiose no ciclo de vida desses organismos.
Em animais multicelulares, G. ocorre em órgãos especiais - as gônadas ou gônadas (ovários, testículos e gônadas hermafroditas) e consiste em três estágios principais: 1) a reprodução das células germinativas primárias - gametogonia (espermatogônias e oogônias) até uma série de mitoses sucessivas. 2) o crescimento e a maturação dessas células são agora chamados de gametócitos (espermatócitos e oócitos), que, como a gametogonia, possuem um conjunto completo (principalmente diplóide) de cromossomos (ver Cromossomos). Neste momento, ocorre o evento principal de G. em animais - a divisão dos gametócitos por meiose, levando a uma redução (redução pela metade) do número de cromossomos nessas células e sua transformação em células haplóides (ver Haploides) - espermátides e ootídeos ; 3) a formação de espermatozoides (ou espermatozoides) e óvulos; neste caso, os ovos são revestidos com uma série de membranas embrionárias, e os espermatozóides adquirem flagelos, que garantem sua mobilidade. Em mulheres de muitas espécies animais, a meiose e a formação do óvulo são concluídas após a penetração do esperma no citoplasma do ovócito, mas antes da fusão dos núcleos do espermatozóide e do óvulo.
Nas plantas, G. é separado da meiose e começa nas células haplóides, nos esporos (nas plantas superiores, micrósporos e megásporos). A partir dos esporos, a geração sexual da planta se desenvolve - o gametófito haplóide, nos genitais dos quais - gametangia (masculino - anteridia (ver Anteridium), feminino - archegonia (Ver Archegonia)) por mitose, G. A exceção são as gimnospermas e angiospermas, nas quais a espermatogênese vai diretamente para o micrósporo germinativo - a célula de pólen. Em todas as plantas de esporos inferiores e superiores, G. em antheridia é uma divisão múltipla de células, como resultado da qual um grande número de pequenos espermatozóides móveis são formados. G. em arquegônia - a formação de um, dois ou mais oócitos. Nas gimnospermas e angiospermas, G. masculino consiste em dividir (por mitose) o núcleo da célula polínica em generativa e vegetativa e posteriormente dividir (também por mitose) do núcleo generativo em dois espermatozoides. Essa divisão ocorre no tubo polínico em germinação. G. fêmea em angiospermas é o isolamento por mitose de uma célula-ovo dentro de um saco embrionário de oito núcleos. Principal diferença de G. em animais e plantas: em animais, combina a transformação de células diplóides em haplóides e a formação de gametas haplóides; nas plantas, G. é reduzido à formação de gametas a partir de células haplóides.
№82 Cariótipo - um conjunto de características (número, tamanho, forma, etc.) de um conjunto completo de cromossomos inerentes às células de uma determinada espécie biológica (cariótipo da espécie), um determinado organismo (cariótipo individual) ou linha (clone) de células. A representação visual de um conjunto completo de cromossomos (cariograma) às vezes também é chamada de cariótipo. O aparecimento dos cromossomos muda significativamente durante o ciclo celular: durante a interfase, os cromossomos ficam localizados no núcleo, via de regra, despiralizados e de difícil observação, portanto, as células são utilizadas para determinar o cariótipo em uma das etapas de sua divisão - mitose metáfase. Para o procedimento de determinação do cariótipo, qualquer população de células em divisão pode ser usada. Para determinar o cariótipo humano, via de regra, utilizam-se linfócitos do sangue periférico, cuja transição do estágio de repouso G0 para a proliferação é provocada pela adição de mitogenafitohemaglutinina. As células da medula óssea ou a cultura primária de fibroblastos da pele também podem ser usadas para determinar o cariótipo. Para aumentar o número de células na fase de metáfase, a colchicinilinocadazole é adicionada à cultura celular pouco antes da fixação, o que bloqueia a formação dos microtúbulos, evitando assim que as cromátides divergem para os pólos de divisão celular e a conclusão da mitose.
Após a fixação, as preparações dos cromossomos metafásicos são coradas e fotografadas; o chamado cariótipo sistemático é formado por micrografias - um conjunto numerado de pares de cromossomos homólogos, as imagens dos cromossomos são orientadas verticalmente com os braços curtos para cima, sua numeração é realizada em ordem decrescente de tamanho, um par de cromossomos sexuais é colocado em o final do conjunto (ver Fig. 1).
Historicamente, os primeiros cariótipos não detalhados que permitiam a classificação de acordo com a morfologia dos cromossomos eram obtidos pela coloração de Romanovsky-Giemsa, porém, maiores detalhamentos da estrutura dos cromossomos nos cariótipos tornaram-se possíveis com o advento das técnicas de coloração cromossômica diferencial. A técnica mais comumente usada em genética médica é a coloração do cromossomo diferencial G.
A comparação dos complexos de marcas transversais no cariótipo clássico ou em regiões com características espectrais específicas permite identificar tanto os cromossomos homólogos quanto suas regiões individuais, o que permite determinar em detalhes as aberrações cromossômicas - rearranjos intra e intercromossômicos, acompanhados de violação da ordem dos fragmentos cromossômicos (deleções, duplicações, inversões, translocações). Tal análise é de grande importância na prática médica, permitindo o diagnóstico de uma série de doenças cromossômicas causadas por violações grosseiras de cariótipos (anormalidades no número de cromossomos) e uma violação da estrutura cromossômica ou multiplicidade de cariótipos celulares no corpo (mosaicismo).
№83 meiose- divisão de redução, divisão de maturação, um método de divisão celular, como resultado do qual há uma diminuição (redução) no número de cromossomos pela metade e uma célula diplóide (contendo dois conjuntos de cromossomos) após dois, rapidamente um após o outro divisões dá origem a 4 haplóides (contendo um conjunto de cromossomos).
Valor biológico M ... é manter a consistênciaCariótipo e em várias gerações de organismos desse tipo e garantindo a possibilidade de recombinação de cromossomos e genes durante o processo sexual. M. é um dos principais mecanismos de hereditariedade
A meiose consiste em 2 divisões consecutivas com uma curta interfase entre elas.
Prófase I - a prófase da primeira divisão é muito complexa e consiste em 5 etapas:
Leptoteno ou leptonema - empacotamento de cromossomos, condensação de DNA com a formação de cromossomos na forma de fios finos (os cromossomos são encurtados).
Zigoteno ou zigoneme - ocorre conjugação - a combinação de cromossomos homólogos com a formação de estruturas que consistem em dois cromossomos conectados, chamados tétrades ou bivalentes, e sua posterior compactação.
Pachytene ou pachinema - (o estágio mais longo) - em alguns lugares, os cromossomos homólogos estão fortemente conectados, formando quiasmas. Neles existe cruzando- troca de sítios entre cromossomos homólogos.
Diploteno ou diplonema - ocorre a descondensação parcial dos cromossomos, enquanto parte do genoma pode funcionar, ocorrem processos de transcrição (formação de RNA), tradução (síntese de proteínas); cromossomos homólogos permanecem conectados uns aos outros. Em alguns animais, os cromossomos em oócitos neste estágio da prófase meiótica adquirem uma forma característica lâmpada escova cromossomos.
Diacinesia - o DNA se condensa novamente tanto quanto possível, os processos sintéticos param, a casca nuclear se dissolve; centríolos divergem para os pólos; cromossomos homólogos permanecem conectados uns aos outros.
No final da prófase I, os centríolos migram para os pólos da célula, os filamentos do fuso de fissão são formados, a membrana nuclear e os nucléolos são destruídos
Metáfase I - cromossomos bivalentes se alinham ao longo do equador da célula.
Anáfase I - os microtúbulos se contraem, os bivalentes se dividem e os cromossomos divergem para os pólos. É importante notar que, devido à conjugação de cromossomos no zigoteno, cromossomos inteiros divergem para os pólos, cada um consistindo de duas cromátides, e não cromátides separadas, como em mitose.
Telófase I - os cromossomos são despiralizados e um envelope nuclear aparece.
A segunda divisão da meiose segue imediatamente a primeira, sem uma interfase pronunciada: o período S está ausente, uma vez que não há replicação de DNA antes da segunda divisão.
Prófase II - ocorre a condensação dos cromossomos, o centro da célula se divide e os produtos de sua divisão divergem para os pólos do núcleo, o envelope nuclear é destruído, um fuso de fissão é formado.
Metáfase II - cromossomos univalentes (cada um consistindo de duas cromátides) estão localizados no "equador" (a uma distância igual dos "pólos" do núcleo) em um plano, formando a chamada placa metafásica.
Anáfase II - univalentes dividem ecromátidesdivergem para os pólos.
Telófase II - os cromossomos são despiralizados e um envelope nuclear aparece.
Como resultado, de umcélula diplóidequatrocélulas haplóides... Nos casos em que a meiose está associada comgametogênese(por exemplo, em animais multicelulares), durante o desenvolvimentooócitos a primeira e a segunda divisões da meiose são agudamente desiguais. Como resultado, um ovo haplóide e três os chamados corpos de redução(derivadas abortivas da primeira e segunda divisões).
№84 Divisão celular - mitose
Se você observasse a divisão celular ao microscópio, veria o seguinte. Primeiro, o material dentro do núcleo se torna muito denso. Você pode até distinguir entre estruturas discretas - cromossomos. Você sabe que as células somáticas humanas (não sexuais) têm 23 pares de cromossomos - um cromossomo de cada par de cada pai. No total, existem 46 cromossomos nas células somáticas humanas. Todos os 46 se alinham no plano equatorial da célula. Nesse estágio, é claramente visível que cada um dos 46 cromossomos, na verdade, consiste em duas cromátides, conectadas entre si em uma região semelhante a um botão. Esta área foi chamada de centrômero. Nas extremidades opostas da célula, existem dois pontos densos (centríolos), cada um dos quais forma os fios do fuso de divisão. Cada fita se junta ao cromossomo no centrômero. Duas fitas são anexadas a cada par de cromossomos - uma de cada centríolo. Então veríamos que os centríolos, como espirais, enrolam os fios do fuso de fissão, como resultado do qual os cromossomos divergem para os pólos da célula. Em seguida, o conteúdo da célula é dividido. Esta é uma visão incrível.
Os eventos se desenvolvem de acordo com o seguinte cenário. É preciso entender que para a divisão celular uma tarefa muito importante deve ser resolvida - a duplicação (replicação) do DNA deve ocorrer, pois é no DNA que estão codificadas todas as informações sobre o que e como deve acontecer na célula. Lembre-se de que o DNA é uma dupla hélice. Fora da espiral está a espinha dorsal do açúcar-fosfato, na qual os açúcares estão ligados uns aos outros por ligações covalentes através do resíduo de ácido fosfórico. Dentro da espiral existem bases nitrogenadas orgânicas conectadas aos pares por pontes de hidrogênio. O significado de copiar informações armazenadas em um cromossomo e transferir essas informações para uma nova célula sem mudanças é muito simples: é necessário desenrolar a dupla hélice do DNA e quebrar as ligações de hidrogênio que mantêm os pares de bases nitrogenadas juntos. A partir desse momento, aparecem duas cadeias simples com bases nitrogenadas, projetando-se da estrutura açúcar-fosfato. Lembre-se de que cada base nitrogenada de uma das fitas de DNA sempre forma ligações com uma base muito específica da outra fita de DNA. Assim, a adenina liga-se apenas à timina e a guanina apenas à citosina. Este é o princípio da complementaridade das bases nitrogenadas. Lembre-se também que os nucleotídeos são moléculas compostas por uma base orgânica e açúcar fosforilado. Em teoria, se os nucleotídeos necessários estiverem presentes no nucleoplasma, cada base de nitrogênio de cada fita de DNA corresponderá a um par adequado. Existe uma enzima específica e muito importante, a DNA polimerase, que torna essa reação possível. Os açúcares da fita de DNA filha recém-formada são conectados uns aos outros por meio de ligações fosfodiéster, como resultado da formação de um esqueleto açúcar-fosfato - e assim aconteceu! Você tem dois cromossomos idênticos onde tinha apenas um (Figura 2.7). É claro que, de fato, o processo de replicação de uma molécula de DNA em uma célula é muito mais complicado e depende do funcionamento de várias enzimas, mas falaremos sobre isso a seguir. Agora vamos dar uma olhada mais de perto no processo de divisão celular. Inicialmente, os pares de cromossomos estão em um estado despiralizado no núcleo e são indistinguíveis ao microscópio. Diz-se que essas células estão em interfase. Nestes, o material genético conhecido como cromatina é uma massa emaranhada e não é tão compacta como quando os cromossomos são distinguíveis ao microscópio. A cromatina deve seu nome ao fato de ser tingida de escuro com tintas de cores especiais. Isso aconteceu muito antes que as pessoas soubessem de sua verdadeira função. Como já mencionado, as células somáticas humanas têm 23 pares de cromossomos. Vinte e dois pares são chamados de cromossomos autossômicos, que são iguais em homens e mulheres. O vigésimo terceiro par é representado por cromossomos sexuais, que são diferentes em homens e mulheres. As mulheres têm dois grandes cromossomos chamados cromossomos X, enquanto os homens têm um cromossomo X grande e um cromossomo Y pequeno. Os cromossomos de um par têm a mesma estrutura. No entanto, os cromossomos recebidos de pais diferentes carregam informações diferentes e, nesse sentido, os pares de cromossomos não são idênticos.
A mitose mostrada na Fig. 2.8, é uma forma universal de dividir qualquer célula eucariótica. A mitose consiste em quatro fases distintas principais: prófase, metáfase, anáfase, telófase. Quando uma célula começa a se dividir, o material genético de cada cromossomo já dobrou. Na prófase, o número de cromossomos corresponde a 46, mas cada cromossomo é duplo, ou seja, é composto por duas cromátides irmãs.
Para começar a duplicar o material genético, a célula precisa resolver uma tarefa difícil - desenrolar a hélice do DNA. A maioria de nós acha que o cromossomo não é tão longo, porque é assim que estamos acostumados a vê-los em todos os desenhos. Na realidade, em uma célula em interfase, a molécula de DNA está em um estado superenrolado, ou seja, está torcida com muita força e em uma determinada ordem. Indicativo é o fato de que se a molécula de DNA estivesse dentro da célula sem torção, seu comprimento seria de cerca de 1 m! Imagine como deve ser complexo e ordenado o empacotamento de tanto material genético dentro de uma célula microscópica. Portanto, fica claro por que, nas células eucarióticas, a dupla hélice da molécula de DNA é compactada em uma estrutura compacta formada pela interação com uma série de proteínas chamadas histonas. Assim, as proteínas cromossômicas empacotam a molécula gigante em um núcleo com um volume de apenas alguns mícrons cúbicos (Fig. 2.9) Imediatamente antes do início do processo de replicação, a molécula de DNA é desenrolada pela enzima helicase. Em seguida, as ligações de hidrogênio são quebradas, as cadeias divergem e as cadeias filhas são formadas em sua matriz de acordo com o princípio da complementaridade.A divisão celular é estritamente controlada. Existem células que se dividem com frequência. Isso inclui células da medula óssea. Outras células, como as células ósseas, dividem-se principalmente durante o crescimento ou dano de um órgão ativo. Finalmente, existem células que se dividem muito raramente ou nunca se dividem. Este tipo inclui células nervosas maduras. O crescimento e a divisão celular descontrolados são um sinal de sua degeneração cancerosa. O processo de divisão celular requer reguladores chamados fatores de ativação. Fatores de crescimento são exemplos de tais fábricas. O estudo dos fatores de crescimento é uma das áreas de crescimento mais rápido na interseção da biologia moderna e da medicina. Muitos fatores estão atualmente destacados e parcialmente caracterizados.
Os fatores de ativação geralmente atuam alterando a forma do DNA e permitindo que a DNA polimerase, que é responsável pela replicação do DNA, se ligue a ele. A DNA polimerase encontra primeiro um local específico na molécula de DNA. Esta região em forma de alça é necessária para que a DNA polimerase se ligue ao DNA. Por sua vez, é formado quando o fator de ativação se liga ao DNA. O local da molécula de DNA onde a DNA polimerase é anexada e o processo de replicação começa é chamado de ponto de origem da replicação. Você também deve estar ciente de que sequências curtas de RNA são necessárias para iniciar a síntese de DNA, que funcionam como “primers”. Tendo se ligado a uma molécula de DNA, a DNA polimerase se move ao longo da cadeia materna, completando a construção da cadeia filha de acordo com o princípio da complementaridade.
Uma das funções mais importantes da DNA polimerase é corrigir erros na fita de DNA recém-sintetizada. Reconhecendo os pares de bases errados, a DNA polimerase volta um passo, a base errada é removida da fita filha e a correta é inserida. Como resultado, o número de erros na replicação do DNA é aproximadamente um erro em 10-100 bilhões de pares de bases. No entanto, como as células se dividem com frequência suficiente, os erros inevitavelmente se acumulam. Muitos desses erros afetam a viabilidade celular. Se as células nas quais ocorreu a mutação sobrevivem, o sistema imunológico as destrói com a ajuda de células especiais adaptadas para esse fim.
Deixe-me voltar agora ao processo de divisão celular. Na prófase da mitose, os cromossomos consistem em duas cromátides, que é o resultado da replicação do DNA na interfase. No entanto, não é possível registrá-los ao microscópio, uma vez que na prófase as cromátides irmãs estão em contato próximo umas com as outras, espiralando-se mutuamente uma em relação à outra. Na prometáfase, a membrana nuclear é destruída e os cromossomos começam a se mover em direção ao plano equatorial da célula. Esse processo garante a separação normal dos cromossomos para os pólos da célula durante a divisão celular, como resultado da qual um conjunto completo de cromossomos entra em cada uma das células recém-formadas. As cromátides irmãs de um cromossomo estão ligadas entre si em um ponto denominado centrômero. Na área do centrômero, existe um cinetocoro, uma estrutura protéica complexa associada ao DNA da região centromérica. Os filamentos do fuso de fissão são fixados a esta zona durante a mitose. Como você já observou, os centríolos estão localizados nos pólos da célula, cujo número também dobra antes da divisão celular. Os centríolos estão envolvidos na formação dos filamentos do fuso de fissão. Cada filamento do fuso se liga a um cromossomo na região do cinetocoro. Na segunda fase da mitose, a metáfase, os cromossomos, constituídos por duas cromátides, se alinham no plano equatorial da célula.
Os filamentos do fuso de fissão puxam os cromossomos, chamados a partir deste momento de cromossomos filhos, para extremidades opostas da célula. A fase durante a qual os cromossomos se movem em direções diferentes é chamada de anáfase. Se tudo correr conforme o planejado, o centríolo se divide e as cromátides irmãs divergem para os pólos da célula. Se os cromossomos não se alinham no plano equatorial da célula na ordem correta, o processo de divisão celular pode ser interrompido.
Depois que os cromossomos se espalharam para extremidades opostas da célula-mãe, a célula se divide em duas como resultado da citocinese. O quarto e último estágio da mitose é denominado telófase. As células que se multiplicam por mitose, como já mencionado, são chamadas de somáticas. Isso inclui todas as células do nosso corpo, com exceção das células que mais tarde se transformam em espermatozoides ou óvulos. As células que contribuem para a formação de um novo organismo são chamadas de células germinativas. As células sexuais são formadas como resultado da meiose, que consideraremos na próxima seção.
A mitose geralmente se resolve sem demora. No entanto, a qualquer momento, milhões de divisões celulares ocorrem no corpo humano. De vez em quando, há muito material genético nas células, enquanto em outras pode ser menos do que o normal. Normalmente, essas células não sobrevivem ou são removidas pelo sistema imunológico.
№85 Doenças hereditárias. Classificação das doenças hereditárias. Doenças hereditárias - doenças cuja ocorrência e desenvolvimento estão associados a defeitos no software das células, herdados através dos gametas. O termo é usado em relação às doenças polietiológicas, em contraste com um grupo mais restrito - as doenças genéticas.
Dependendo do tipo de células afetadas primárias, os seguintes grupos de doenças são identificados: doenças devido a mutações nas células germinativas - "gaméticas", ou seja, doenças realmente hereditárias (por exemplo, fenilcetonúria, hemofilia); essas doenças são hereditárias; doenças devido a mutações em células somáticas - "somáticas" (por exemplo, tumores, algumas doenças de autoagressão imunológica). Essas doenças não são hereditárias; doenças devido a uma combinação de mutações em células germinativas e somáticas (por exemplo, retinoblastoma familiar).
As doenças letais, subletais e hipogenitais são consideradas separadamente: as doenças letais levam à morte durante o desenvolvimento intra-uterino (por exemplo, monossomia por autossomos, haploidia, a maioria da poliploidia); doenças subletais levam à morte do indivíduo antes da puberdade (por exemplo, imunodeficiências hereditárias, como agam-maglobulinemia do tipo Swiss, síndrome de Louis-Bar, alguma hemofilia); doenças hipogenitais são combinadas com infertilidade (por exemplo, Shereshevsky-Turner, síndromes de Kleinfelter).
Dependendo do papel da hereditariedade e do ambiente, quatro grupos de doenças são distinguidos: doenças hereditárias propriamente ditas (desenvolvem-se como resultado de mutações); doenças ecogenéticas (se desenvolvem na presença de uma mutação sob a influência de um ambiente de fator específico); doenças de predisposição genética (a contribuição dos fatores genéticos é óbvia); doenças causadas por fatores ambientais (a rigor, este grupo de doenças não pertence às doenças hereditárias, pois a contribuição dos fatores genéticos para a sua ocorrência, desenvolvimento e desfecho pode ser extremamente pequena, duvidosa ou desconhecida).