Mendel tegeles herneste valikuga ning just herneste, teaduslikule õnnele ja Mendeli katsete rangusele võlgneme põhiliste pärimisseaduste avastamise: esimese põlvkonna hübriidide ühetaolisuse seaduse, poolitamise seaduse ja sõltumatu kombinatsiooni seadus.
Mendel katsetas 22 hernesordiga, mis erinevad üksteisest 7 omaduse (värvus, seemne tekstuur) poolest. Mendel juhtis oma tööd kaheksa aastat, uuris 10 000 hernetaime. Kõik herned, mida ta uuris, olid puhtast liinist; selliste taimede omavahelise ristamise tulemused on alati olnud samad. Mendel tutvustas oma töö tulemusi 1865. aastal artiklis, millest sai geneetika nurgakivi. Raske on öelda, mis temas ja tema töös rohkem imetlust väärib - katsete rangus, tulemuste esituse selgus, katsematerjali täiuslik tundmine või eelkäijate töö tundmine.
Mendeli kolleegid ja kaasaegsed ei suutnud hinnata tema järelduste tähtsust. Vastavalt A.E. Gaisinovitš, kuni 19. sajandi lõpuni. seda tsiteeriti vaid viis korda ja ainult üks teadlane - vene botaanik I.O. Schmalhausen – hindas selle töö tähtsust. Kuid 20. sajandi alguses avastasid tema avastatud seadused peaaegu samaaegselt ja üksteisest sõltumatult teadlased K. Correns, E. Cermak ja K. de Vries. Nende avastuste tähtsus sai teadusringkondadele kohe selgeks 1900. aastate alguses; nende äratundmist seostati teatud eduga tsütoloogias ja tuumapärilikkuse hüpoteesi kujunemisega.
|
Tunnuste pärimise peamised mustrid. Inimese pärilikkust on uuritud palju vähemal määral kui taime- ja loomaorganismide tunnuste pärimise mehhanisme. Sellegipoolest on juba saadud huvitavaid andmeid inimese paljude füüsiliste omaduste päriliku sõltuvuse kohta. Kaasaegne geeniteaduse tase lubab väita, et kõik taimede ja loomadega tehtud katsetes tuvastatud tunnuste pärilikkuse põhimustrid ja pärilikkuse seadused kehtivad ka inimeste puhul. Näiteks kui noormees, kellel on mõlemal homoloogsel kromosoomil pruunide silmade geenid (homosügootsed pruunid), abiellub sinisilmse tüdrukuga, kelle kromosoomid sisaldavad samuti ainult siniste silmade geene, siis pärivad nende järglased avastatud seaduste järgi silmade värvi. geneetika rajaja G. Mendeli poolt eelmise sajandi 60ndatel oma katsetes herneste hübriididega. Inimestel ja enamikul loomadel toimub ka sooline pärand vastavalt üldistele seadustele ja on seotud kromosoomide jaotumisega, millel on X- ja K-kujuline vorm (sugukromosoomid). Naise keha kromosoomikomplekt sisaldab kahte X-kromosoomi ja mehe kehas on üks X-kromosoom ja üks Y-kromosoom (vt joonis 7). Selliste näidete hulk võib olla lõpmatult suur ja pole kahtlustki, et inimese “geeniportree” (genotüüp) määrab suuresti ära paljud tema välised omadused (fenotüüp). Allpool on loetletud mõned inimese pärilikud tunnused, mille määravad domineerivad ja retsessiivsed geenid (K-Willie, 1974 järgi). Geenimõju avaldumist saab läbi viia erinevatel ontogeneesi etappidel, kuid enamik fenotüüpilisi tunnuseid määratakse juba enne sündi. Lõpuks ei ole geenide ilmnemine surmav, vaid sõltub keskkonnateguritest. Näiteks ei arene välja raske pärilik haigus fenüülketonuuria, kui laps, kelle genotüübis on seda haigust põhjustavaid genotüüpe, hakkab kohe pärast sündi teatud dieeti saama. Seega on geneetikal veenvaid fakte, mis tõendavad nii rangelt pärilike tunnuste (näiteks veretase, hemofiilia, juuste ja silmade värv, näojooned ja paljud teised) kui ka väliskeskkonnast suuresti määratud tunnuste olemasolu (nt. inimese kasv ja mass, tema lihaste tugevus ja osavus, kalduvus haigustele jne). Tuleb märkida, et geenide ja tunnuste vahel puudub otsene seos: ühe tunnuse areng võib sõltuda paljude geenide mõjust ja üks geen võib mõjutada paljude tunnuste kujunemist. Seda pilti muudab veelgi keerulisemaks pidev väliskeskkonnast tulenev korrektsioon. Järelikult on rahvastiku omand ja areng alati omavahel tihedalt seotud. Lapse arengut on võimatu ette kujutada ilma keskkonna korrigeerivate mõjudeta, nagu on võimatu ilma geneetiliselt etteantud arenguprogrammita, mis on süntees kõigi eelnevate põlvkondade ajaloolisest kogemusest. Mõelgem pärilikkuse ja keskkonna rollile inimese mõningate füüsiliste omaduste kujunemisel tema sünnieelses ja -järgses arengus. Sünnieelne areng. Lapse elundite ja funktsionaalsete süsteemide moodustumine embrüogeneesi protsessis on genotüübi kontrolli all, kuid keskkonnategurid mängivad olulist rolli. Embrüo jaoks on ema organism esmane keskkond. Siin, spetsiaalses elundis - emakas - on embrüo suhteliselt hästi kaitstud paljude kahjulike mõjude eest ja saab platsenta kaudu kõik oma olemasoluks vajaliku. Sellegipoolest, eriti arengu algstaadiumis, mõjutavad embrüo arengut ka paljud ema keha mõjutavad tegurid (kõige olulisemad on: ioniseeriv kiirgus, naise raseduse ajal põdevad haigused ja paljud kemikaalid: alkohol, nikotiin, antibiootikumid, hormonaalsed ravimid jne). Tuleb märkida, et inimese sünnieelsel perioodil on kriitilisi perioode, mil arenev organism on välistegurite toime suhtes eriti tundlik. Selliseid perioode on kaks. Esimene hõlmab sünnieelse arengu algust, selle esimest kolme nädalat. Sel ajal toimub kõigi kõige olulisemate elundite munemine ja selle perioodi kahjulikud mõjud põhjustavad sageli embrüo surma. Teisel kriitilisel perioodil (4. kuni 7. nädal) toimub kõigi organite edasine areng ning sellel perioodil võivad kahjulikud välismõjud põhjustada erinevate füüsiliste defektidega lapse sündi (joon. 9). On oluline, et samu füüsilisi sünnidefekte saaks seostada geneetiliste struktuuride kahjustusega ja ebasoodsate välistegurite toimega embrüogeneesi protsessis. See on hea tõend selle kohta, et keskkond ja pärilikkus vastutavad lapse normaalse arengu eest võrdselt. Seega on sünnihetkeks üldiselt moodustunud kõik inimese elundid ja tema füsioloogilised süsteemid, sealhulgas närvisüsteem. Loomulikult tekib küsimus: kas inimese vaimse tegevuse aluseks olevad närvisüsteemi omadused, tema mõistuse alus, on pärilikult ette määratud? Kas meie keha eluprotsesside reguleerimisega seotud madalama närvitegevuse jaoks on olemas valmis morfoloogilisi "ettevalmistusi teadvuseks"? Nendele küsimustele vastamiseks peatume inimese arengu järgmisel, sünnijärgsel etapil. Lapse sünnijärgne areng. Kõigepealt käsitleme peamisi meetodeid, mis võimaldavad uurida keskkonna ja pärilikkuse spetsiifilist tähtsust laste ja noorukite sünnijärgses arengus. Selliste katsete läbiviimine taimedega on lihtne ja seisneb kahe genotüübilt identse organismirühma eraldamises ja nende rühmade paigutamises erinevatesse elutingimustesse. Näiteks üht taimerühma (kontroll) kasvatatakse tavatingimustes, teist (katselist) - pimendatud ruumis. Selliste katsete tulemusena võib järeldada, et rohelise klorofülli teke taimedes ei sõltu ainult pärilikkusest, vaid ka keskkonnateguritest (valgus), kuna pimedas kasvatatud taimed seda pigmenti ei sisalda. Sarnaste katsete läbiviimine inimestega on võimatu nii moraalsest ja eetilisest kui ka bioloogilisest seisukohast. Maailmas pole kahte ühesuguse genotüübiga inimest. Kuid ... reeglist on erandeid. Need on identsed või homosügootsed kaksikud (IB), millel on mitte ainult silmatorkav välised sarnasused, vaid ka peaaegu identsed geeniportreed. Identsete ja mitteidentsete kaksikute (TW) sünd pole haruldane, keskmiselt lõpeb üks 100-st rasedusest rohkem kui ühe lapse sünniga. Kaksikud on kõige suurepärasem "materjal", mille loodus ise teadlasele esitab, seetõttu nimetatakse meetodit ennast kaksikuks. Mis on selle olemus? Oletame, et peate välja selgitama, milline tegur on lapse füüsilise arengu ja kasvu eest kõige enam vastutav: pärilikkus või keskkond? Selleks selgitatakse välja kaksikpaarid, tehakse nende morfoloogiline uuring ja uuritakse elustiili (tavaliselt on kaksikud, eriti identsed, sarnased huvid ja lähedased kasvatustingimused). Läbiviidud uuringute põhjal tehakse kindlaks kaksikute sarnasuse koefitsient (konkordantsus) ning tehakse järeldus pärilikkuse või keskkonna rollist teatud omaduste kujunemisel. Selliste identsete ja mitteidentsete kaksikute uuringute tulemused mõne haiguse esinemissageduse osas on esitatud tabelis. Kahjuks teevad õpetajad andekate laste väljaselgitamiseks endiselt vähe tööd, mida sageli seostatakse laste füsioloogilise ja vaimse individuaalsuse alahindamise ning lapse elementaarse bioloogia teadmatusega. Nõrk töö laste andekuse väljaselgitamisel on seletatav ka selle valdkonna endiselt vähese uurimistööga ja täpsete teaduslike kriteeriumide puudumisega, mille järgi oleks õpetajal lihtne lapse pärilikke kalduvusi tuvastada. Andekuse ja närvisüsteemi kõrge erutuvuse vahel on vaid tihe seos, mille sümptomiks on sageli lapse terav tasakaalutus (psühhopaatia). Õpetaja peaks selliste laste suhtes olema eriti tähelepanelik. See on vajalik eelkõige hügieenilisest küljest, kuna õpetaja vale tegevus võib viia äärmuslike iseloomuomaduste kujunemiseni patoloogilisteks. See on vajalik ka pärilike kalduvuste õigeaegseks tuvastamiseks ja nende optimaalseks arenguks. Oluline on meeles pidada, et ilmne õppimishimu puudumine, väline laiskus ja distsiplineerimatus ei ole piisavad tõendid võimete puudumise kohta. Ajaloost on palju tõendeid selle järelduse kohta. Relatiivsusteooria looja A. Einstein lapsepõlves avatud võimeid ei näidanud ja alles 9-aastaselt suutis minna ettevalmistuskooli. A. Humboldt (1769-1859) ja L. Pasteur (1822-1895), silmapaistev inglise füüsik I. Newton, olid koolis tuntud laiska ja saamatu õpilasena. Võib arvata, et vanemate ja õpetajate oskuslik kohtlemine neisse lõi lõpuks kõik vajalikud tingimused annete arendamiseks. Pealegi on geniaalsuse kõige olulisem omadus raske töö. Suur Ameerika leiutaja T. Edison kirjutas: "Geenius on üks protsent inspiratsioonist ja üheksakümmend üheksa protsenti higistamine." See, kes ei õpi "higistama", ei suuda hoolimata kõige silmapaistvamatest pärilikest kalduvustest kunagi saavutada suurt edu üheski inimtegevuses. Kes, kui mitte õpetaja, peaks ennekõike kasvatama lapses töökust - see on isiksuse järkjärgulise arengu kõige olulisem tegur, mis tahes võimete kujunemise peamine tingimus. Seega pärilikkus annab ainult "toorainet" ja keskkond teostab oma peamist "töötlemist". Inimene, kes on sündinud ka kõige soodsamate kalduvustega, kuid elab keskkonnas, mis takistab tema võimete arengut, jääb keskpärasuseks, nii et peamine vastutus lapse intellektuaalsete võimete arendamise eest langeb kasvatajatele. Sellegipoolest ei tohiks absolutiseerida keskkonna rolli, ei tohi unustada, et pärilikkusest saadav “materjal” ei ole alati kvaliteetne ning ka kõige andekam skulptor ei suuda liivast skulptuuri voolida, nagu marmorplokk iseenesest ei muutu kunstiteoseks ... №67 - nr 68. Autosoomne domineeriv pärand- pärilikkuse tüüp, mille puhul piisab haiguse (või tunnuse) avaldumiseks ühest autosoomi lokaliseeritud mutantse alleelist. Autosoomne retsessiivne pärand- tunnuse või haiguse pärilikkuse tüüp, mille puhul autosoomis lokaliseeritud mutantne alleel peab olema päritud mõlemalt vanemalt. |
№69. Sooga seotud pärand - sugukromosoomides paikneva geeni pärand. Tunnuste pärandumist, mis ilmnevad ainult samast soost isenditel, kuid mida ei määra sugukromosoomides paiknevad geenid, nimetatakse soopiiranguks.
X-kromosoomiga seotud pärand on geenide pärand, kui meessugu on heterogameetiline ja seda iseloomustab Y-kromosoomi (XY) olemasolu ning naised on homogameetilised ja neil on kaks X-kromosoomi (XX). Seda tüüpi pärilikkus on kõigil imetajatel (kaasa arvatud inimestel), enamikul putukatel ja roomajatel.
№70 Hollandi pärandtüüp. Holandri pärand (holandricinheritance) – Y-kromosoomi mittehomoloogses osas paikneva geeni poolt kontrollitava tunnuse pärand. Holandrikgeen on geen, mis on seotud meessoo Y-kromosoomiga. G. g määratud märgid kanduvad edasi ainult meesliini kaudu – isalt pojale.
№71 X-seotud pärimistüübiga mutantne geen asub X-kromosoomis. Kui mutatsioonil on domineeriv mõju, võivad haiged olla nii mehed kui naised. Haigus kandub aga haigelt isalt 100% tõenäosusega edasi ainult tüdrukutele, aga mitte poistele, kes saavad isalt Y-kromosoomi (joon. 5). Domineeriva X-seotud mutatsiooni ülekandumise tõenäosus haigelt emalt lastele on 50%. Ühtviisi suure tõenäosusega pärivad haigust nii tütar kui ka poeg.
Domineerivale,seotud X-kromosoomiga, pärilikkuse tüübiks on lastearstidele tuntud patoloogia, D-vitamiini suhtes resistentne rahhiit (sünonüümid: hüpofosfateemia, perekondlik X-seotud hüpofosfateemia, fosfaatdiabeet). Selle raskekujulise rahhiidi diagnoosimist, mis ei kao suurte D-vitamiini annuste mõjul, kinnitab sarnase haiguse esinemine mõnel sugulasel, nii meestel kui naistel.
Sagedamini X-seotud haigusedon päritud retsessiivselt. Retsessiivsete haiguste eripäraX-seotud pärimise tüüp on see, et mehed on peres haiged ja nad pärivad mutantse alleeli oma praktiliselt tervelt emalt, mis on mutantse alleeli suhtes heterosügootne. Tõuraamatu koostamisel on sellistel emadel sageli haiged vennad või onud. Haiged mehed võivad oma haigust edasi anda ainult ühe põlvkonna jooksul ja ainult lastelastele (kuid mitte lapselastele) oma terve, kuid heterosügootse tütre kaudu.
Seega, kui jälitate sugupuu pärandit meesliinisretsessiivne X-seotud haigus, saate midagi "malerüütli käigu" sarnast.
Enamikteadaolevad X-seotud haigusedonhemofiilia A ja B , samuti lihassüsteemi kõige raskem patoloogia -Duchenne Beckeri lihasdüstroofia ... Hemofiilia A areng põhineb vere hüübimisfaktori VIII sünteesi eest vastutava geeni mutatsioonidel ja hemofiilia B korral on vere hüübimisfaktor IX defektne. Mõlemad VIII ja IX faktorit kodeerivad geenid paiknevad X-kromosoomi pikas harus vastavalt q28 ja q27.1-2 juures. On teada, et hemofiilia korral on vere hüübimise rikkumine ja väikseimad lõiked võivad ilma erilise hematoloogilise hoolduseta patsiendi surma viia. Pange tähele, et naistel - hemofiilia geeni kandjatel (nn "juhid") on mõnel juhul ka kalduvus verejooksule, mis väljendub raskes menstruatsioonis ja pikaajalises verejooksus sünnituse ajal. Sünnitusarstid-günekoloogid peavad seda asjaolu arvesse võtma, kui nad töötavad naistega, kellel on hemofiilia A või B geenides mutantne alleel.
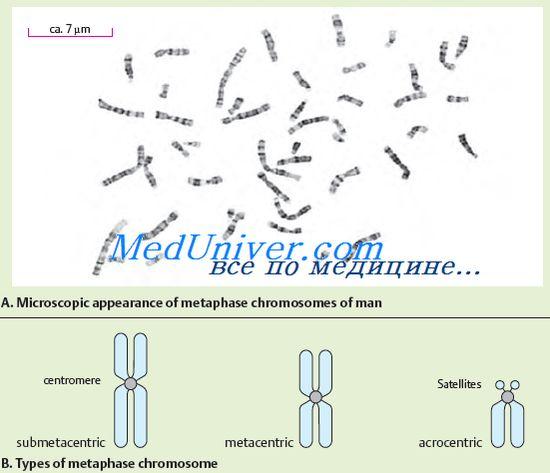
№72 Kromosoomide ehitus ja talitlus. Kromosoomide morfoloogia on kõige paremini nähtav rakus metafaasi staadiumis. Kromosoom koosneb kahest vardakujulisest kehast – kromatiididest. Iga kromosoomi mõlemad kromatiidid on üksteisega geneetiliselt identsed. Kromosoomid on pikkuses erinevad. Kromosoomidel on tsentromeer või esmane ahenemine, kaks telomeeri ja kaks kätt. Mõnel kromosoomil on sekundaarsed kitsendused ja satelliidid isoleeritud. Kromosoomi liikumine määrab tsentromeeri, millel on keeruline struktuur. Tsentromeeri DNA eristub iseloomuliku nukleotiidjärjestuse ja spetsiifiliste valkude poolest. Sõltuvalt tsentromeeri asukohast eristatakse akrotsentrilisi, submetatsentrilisi ja metatsentrilisi kromosoome. Nagu eespool mainitud, on mõnel kromosoomil sekundaarsed kitsendused. Need, erinevalt primaarsest ahenemisest (tsentromeerist), ei toimi spindli filamentide kinnituskohana ega mängi kromosoomide liikumises mingit rolli. Mõned sekundaarsed kitsendused on seotud nukleoolide moodustumisega, antud juhul nimetatakse neid tuumaorganisaatoriteks. Nukleolaarsed organisaatorid sisaldavad RNA sünteesi eest vastutavaid geene. Teiste sekundaarsete kitsenduste funktsioon pole veel selge. Mõnel akrotsentrilisel kromosoomil on satelliidid – piirkonnad, mis on ülejäänud kromosoomiga ühendatud õhukese kromatiini niidiga. Satelliidi kuju ja suurus on antud kromosoomi puhul konstantsed. Inimestel on satelliidid viies kromosoomipaaris. Struktuurse heterokromatiini rikkaid kromosoomiotsi nimetatakse telomeerideks. Telomeerid takistavad kromosoomide otste kokkukleepumist pärast reduplikatsiooni ja aitavad seeläbi kaasa nende terviklikkuse säilimisele. Järelikult vastutavad telomeerid kromosoomide kui üksikute üksuste olemasolu eest. Kromosoome, millel on sama geenijärjestus, nimetatakse homoloogseteks. Neil on sama struktuur (pikkus, tsentromeeri asukoht jne). Mittehomoloogsetel kromosoomidel on erinev geenikomplekt ja erinev struktuur. Kromosoomide peenstruktuuri uurimine on näidanud, et need koosnevad DNA-st, valgust ja vähesest kogusest RNA-st. DNA molekul kannab endas kogu pikkuses jaotunud negatiivseid laenguid ning sellega seotud valgud – histoonid on positiivse laenguga. Seda DNA-valgu kompleksi nimetatakse kromatiiniks. Kromatiinil võib olla erineva kondenseerumisastmega. Kondenseeritud kromatiini nimetatakse heterokromatiiniks, dekondenseeritud kromatiini nimetatakse eukromatiiniks. Kromatiini dekondensatsiooni aste peegeldab selle funktsionaalset olekut. Heterokromatiini piirkonnad on funktsionaalselt vähem aktiivsed kui eukromatiini piirkonnad, milles enamik geene on lokaliseeritud. Eristage struktuurset heterokromatiini, mille kogus on erinevates kromosoomides erinev, kuid see paikneb pidevalt lähitsentomeersetes piirkondades. Lisaks struktuursele heterokromatiinile on valikuline heterokromatiin, mis ilmub kromosoomi eukromaatiliste piirkondade superkeerdumisel. Selle nähtuse olemasolu inimese kromosoomides kinnitab asjaolu, et naise somaatilistes rakkudes on ühe X-kromosoomi geneetiline inaktiveerimine. Selle olemus seisneb selles, et X-kromosoomis lokaliseeritud geenide teise doosi inaktiveerimiseks on olemas evolutsiooniliselt välja kujunenud mehhanism, mille tulemusena, vaatamata erinevale X-kromosoomide arvule mees- ja naisorganismides, suureneb geenide arv. toimimine neis on võrdne. Kromatiin kondenseerub maksimaalselt mitootilise rakkude jagunemise käigus, siis on seda võimalik tuvastada tihedate kromosoomide kujul.Kromosoomide DNA molekulide mõõtmed on tohutud. Iga kromosoomi esindab üks DNA molekul. Need võivad ulatuda sadade mikromeetrite ja isegi sentimeetriteni. Inimese kromosoomidest on suurim esimene; selle DNA kogupikkus on kuni 7 cm.Ühe inimese raku kõigi kromosoomide DNA molekulide kogupikkus on 170 cm Vaatamata DNA molekulide hiiglaslikule suurusele on ta kromosoomidesse üsna tihedalt pakitud. Selle kromosomaalse DNA spetsiifilise pakkimise pakuvad histooni valgud. Histoonid on paigutatud piki DNA molekuli pikkust plokkide kujul. Üks plokk sisaldab 8 histooni molekuli, mis moodustavad nukleosoomi (moodustis, mis koosneb DNA ahelast, mis on keerdunud ümber histooni oktameeri). Nukleosoomi suurus on umbes 10 nm. Nukleosoomid näevad välja nagu nööri külge kinnitatud helmed. Nukleosoomid ja neid ühendavad DNA lõigud on tihedalt spiraali kujul kokku pakitud, sellise spiraali iga pöörde kohta on kuus nukleosoomi. Nii kujuneb kromosoomi struktuur. Organismi pärilik informatsioon on rangelt järjestatud vastavalt üksikutele kromosoomidele. Iga organismi iseloomustab konkreetne kromosoomide komplekt (arv, suurus ja struktuur), mida nimetatakse karüotüübiks. Inimese karüotüüpi esindab kakskümmend neli erinevat kromosoomi (22 paari autosoome, X- ja Y-kromosoome). Kariotüüp on liigi pass. Karüotüübi analüüsiga saab tuvastada häireid, mis võivad varajases arengujärgus kaasa tuua arenguhäireid, pärilikke haigusi või loote ja embrüote surma. Pikka aega arvati, et inimese karüotüüp koosneb 48 kromosoomist. 1956. aasta alguses avaldati aga raport, mille kohaselt on inimese karüotüübi kromosoomide arv 46. Inimese kromosoomid erinevad suuruse, tsentromeeride asukoha ja sekundaarsete kitsenduste poolest. Esimest korda viidi karüotüübi rühmadesse jagamine läbi 1960. aastal Denveris (USA) toimunud konverentsil. Inimese karüotüübi kirjeldus sisaldas algselt kahte järgmist põhimõtet: kromosoomide paiknemine nende pikkuses; kromosoomide rühmitamine tsentromeeri asukoha järgi (metatsentriline, submetatsentriline, akrotsentriline). Kromosoomide arvu täpne püsivus, individuaalsus ja struktuuri keerukus näitavad nende poolt täidetava funktsiooni tähtsust. Kromosoomid täidavad raku peamise geneetilise aparaadi funktsiooni. Nendes on geenid paigutatud lineaarsesse järjekorda, millest igaüks hõivab kromosoomis rangelt määratletud koha (lookuse). Iga kromosoom sisaldab palju geene, kuid organismi normaalseks arenguks on vaja geenide komplekti täielikust kromosoomikomplektist.
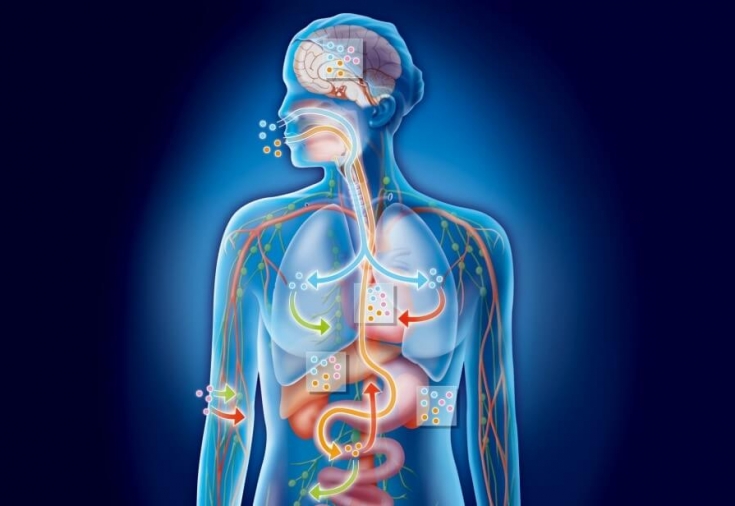
№73. Pärilikkus põhineb kõigi elusorganismide võimel koguda, talletada ja järglastele pärilikku teavet edastada. See elusaine üks olulisemaid kvalitatiivseid omadusi on seotud nukleiinhapetega - desoksüribonukleiinhape (DNA) ja ribonukleiinhape (RNA). Juhtroll kuulub DNA-le - pikima elusorganismide molekulile, mis on koondunud rakkude tuumadesse ja esindab selle pärilikku aparaati. DNA molekuli suur pikkus võimaldab sellele nagu telegraafilindile "üles kirjutada" kõik tulevase organismi põhiomadused ja selle arenguprogramm. Selline "salvestus" toimub spetsiaalse "nukleiinkeele" ehk "nukleiinkoodi" abil, mille sisuks on DNA-d moodustava nelja keemilise ühendi järjestuse muutmine. Piltlikult öeldes koosneb "nukleiinkeel" neljast tähest, millest on ehitatud eraldi sõnad, ja terveid "tuumakeele" lauseid.
Sellisel nukleiinsel "ribal" saab eristada eraldi iseseisvaid alasid, sealhulgas ühe tunnuse arenguprogrammi kirjeldust. Neid nimetatakse geenideks. Iga DNA molekul sisaldab sadu geene ja on programm paljude organismi omaduste ja omaduste arendamiseks. Spetsiaalsete valkude ja mõnede muude ainetega kombineerides moodustavad DNA molekulid tuumas spetsiaalsed moodustised - nn kromosoomid.
Geneetilise materjali organiseerimine. Geenid [kreeka keelest. genos, sünd] – pärilikkuse ühik, DNA tükk, mis hõivab kromosoomis kindla koha. Geneetika seisukohalt on geen pärilik tegur ja geneetilise materjali jagamatu ühik. Struktuurgeen (tsistron) on DNA fragment, mis osaleb polüpeptiidahela moodustamises. See koosneb liiderjärjestusest, kodeerivatest fragmentidest (eksonitest), insertsioonijärjestustest (nitronitest) ja terminaalsest järjestusest. Kuna mõned valgud koosnevad rohkem kui ühest alaühikust, tuleks heteromultimeerse (st koosneb kahest või enamast erinevast polüpeptiidsubühikust) valgu kohta sõnastust "üks geen – üks ensüüm" tõlgendada kui "üks geen - üks polüpeptiidahel". .
Genotüüp on geenide kogum organismis. Isegi iidsetel aegadel kasutasid inimesed empiiriliselt pärimisseadusi. Selle kogemuse põhjal on välja kujunenud valik [al. lat. selectio, select] – teadus uute taime- ja loomatõugude sortide loomise meetodite kohta valiku ja ristamise teel. Kuni viimase ajani tundus genotüüp immutamatu, ei allunud inimtegevusele. Geenide struktuuri avastamine võimaldas neid isoleeritud kujul isoleerida, biokeemiliselt sünteesida ja isegi organismi viia. Sai võimalik geeni mõjutada ilma seda organismist väljutamata. Kõik see lõi eeldused genotüübiga manipuleerimiseks.
№74 Nukleiinhapete struktuur.
Nukleiinhape(ladina keelest nucleus - nucleus) - suure molekulmassiga orgaaniline ühend, nukleotiidide jääkidest moodustunud biopolümeer (polünukleotiid). Nukleiinhapped DNA & RNA esinevad kõigi elusorganismide rakkudes ja täidavad kõige olulisemaid funktsioone päriliku teabe säilitamisel, edastamisel ja rakendamisel. Nukleiinhapete polümeerseid vorme nimetatakse polünukleotiidideks. Nukleotiidide ahelad on ühendatud fosforhappejäägi (fosfodiesterside) kaudu. Kuna nukleotiidides on ainult kahte tüüpi heterotsüklilisi molekule, riboos ja desoksüriboos, on nukleiinhappeid ainult kahte tüüpi - desoksüribonukleiinhape (DNA) ja ribonukleiinhape (RNA).
Monomeerseid vorme leidub ka rakkudes ja neil on oluline roll signaalimisel või energia salvestamisel. Tuntuim RNA monomeer on ATP, adenosiintrifosforhape, raku kõige olulisem energiaakumulaator.
Nukleiinhapete üks peamisi funktsioone on valgusünteesi määramine. Teave DNA nukleotiidjärjestuses kodeeritud valkude struktuuri kohta peab kanduma ühest põlvkonnast teise ja seetõttu tuleb see korrektselt kopeerida, s.t. täpselt sama DNA molekuli süntees (replikatsioon).
№75RepDNA likatsioon desoksüribonukleiinhappe tütarmolekuli sünteesi protsess DNA lähtemolekuli maatriksil. Järgneva emaraku jagunemise käigus saab iga tütarrakk DNA molekuli ühe koopia, mis on identne algse emaraku DNA-ga. See protsess tagab geneetilise teabe täpse edastamise põlvest põlve. DNA replikatsiooni viib läbi kompleksne ensümaatiline kompleks, mis koosneb 15-20 erinevast valgust, mida nimetatakse repisoomiks. DNA replikatsioon on rakkude jagunemise võtmesündmus. On oluline, et jagunemise ajal replitseeritaks DNA täielikult ja ainult üks kord. Seda pakuvad teatud DNA replikatsiooni reguleerimise mehhanismid. Replikatsioon toimub kolmes etapis:
replikatsiooni algatamine
pikenemine
replikatsiooni lõpetamine.
Replikatsiooni reguleerimine toimub peamiselt initsiatsioonifaasis. Seda on üsna lihtne rakendada, sest replikatsioon võib alata mitte mis tahes DNA tükist, vaid rangelt määratletud tükist, mida nimetatakse replikatsiooni initsiatsioonikohaks. Selliseid saite võib genoomis olla ainult üks või mitu. Replikoni mõiste on tihedalt seotud replikatsiooni initsiatsiooni saidi kontseptsiooniga. Replikon on DNA osa, mis sisaldab replikatsiooni initsiatsioonisaiti ja replitseerub, kui DNA süntees sellest kohast algab. Bakterite genoomid esindavad reeglina ühte replikoni, mis tähendab, et kogu genoomi replikatsioon on vaid ühe replikatsiooni initsiatsiooniakti tulemus. Eukarüootide genoomid (nagu ka nende üksikud kromosoomid) koosnevad suurest hulgast sõltumatutest replikonitest, mis vähendab oluliselt üksiku kromosoomi replikatsiooni koguaega. Molekulaarseid mehhanisme, mis kontrollivad replikatsiooni initsiatsiooniaktide arvu igas kohas ühe raku jagunemise tsükli jooksul, nimetatakse koopiate arvu kontrolliks. Lisaks kromosomaalsele DNA-le sisaldavad bakterirakud sageli plasmiide, mis on üksikud replikonid. Plasmiididel on oma koopiakontrolli mehhanismid: nad võivad sünteesida ainult ühe plasmiidi koopia rakutsükli kohta või tuhandeid koopiaid.
Replikatsioon algab replikatsiooni alguse kohast DNA kaksikheeliksi lahtikerimisega, replikatsioonikahvli moodustumisega – DNA otsese replikatsiooni kohaga. Iga sait võib moodustada ühe või kaks replikatsioonikahvlit, olenevalt sellest, kas replikatsioon on ühe- või kahesuunaline. Kahesuunaline replikatsioon on tavalisem. Mõni aeg pärast replikatsiooni algust elektronmikroskoobis on võimalik jälgida replikatsioonisilma – kromosoomi osa, kus DNA on juba replitseeritud ja mida ümbritsevad mittereplitseeruva DNA pikemad lõigud.
Replikatsioonikahvlis kopeerib DNA suurt valgukompleksi (replicisoomi), mille võtmeensüümiks on DNA polümeraas. Replikatsioonikahvel liigub prokarüootidel umbes 100 000 aluspaari minutis ja eukarüootidel 500–5000 aluspaari minutis.
№76 Kromosoomide ehitus ja tüübid.Iga inimkeha somaatilise raku tuum sisaldab46 kromosoomi... komplektkromosoomid iga indiviidi, nii normaalset kui patoloogilist, nimetatakse karüotüübiks.
Alates46 kromosoomiInimese kromosoomikomplekti moodustavad 44 või 22 paari esindavad autosoomikromosoomid , viimane paar on sugukromosoomid. Naistel esindavad sugukromosoomid tavaliselt kahe X-kromosoomiga ja meestel - X- ja Y-kromosoomiga.
Kõikkromosoomide paarid nii autosoomne kui sooline, üks kromosoomidest saadakse isalt ja teine emalt. Ühe paari kromosoome nimetatakse homoloogideks ehk homoloogseteks kromosoomideks. Reproduktiivrakud (sperma ja munad) sisaldavad haploidset kromosoomide komplekti, s.o. 23 kromosoomi.
 Sperma jaguneb kahte tüüpi olenevalt sellest, kas see sisaldabkromosoomX või Y. Tavaliselt sisaldavad kõik munadainult kromosoom
X.
Sperma jaguneb kahte tüüpi olenevalt sellest, kas see sisaldabkromosoomX või Y. Tavaliselt sisaldavad kõik munadainult kromosoom
X.
Kromosoomid on selgelt nähtavad pärast spetsiaalset värvimist rakkude jagunemise ajal, kui kromosoomid on maksimaalselt spiraalitud. Sel juhul tuvastatakse igas kromosoomis ahenemine, mida nimetatakse tsentromeeriks. Tsentromeer jagab kromosoomi lühikeseks käeks (tähistatakse tähega "p") ja pikaks käeks (tähistatakse tähega "q"). Tsentromeer määrab kromosoomi liikumise raku jagunemise ajal. Vastavalt tsentromeeri asukohale liigitatakse kromosoomid mitmesse rühma. Kui tsentromeer asub kromosoomi keskel, siis nimetatakse sellist kromosoomi metatsentriliseks, kui tsentromeer asub kromosoomi ühele otsale lähemal, siis nimetatakse seda akrotsentriliseks. Mõnel akrotsentrilisel kromosoomil on nn satelliidid, mis moodustavad mittejagunevas rakus tuumakesi. Tuumad sisaldavad arvukalt rRNA koopiaid. Lisaks eristatakse submetatsentrilisi kromosoome, kui tsentromeer ei asu kromosoomi keskel, vaid on mõnevõrra nihkunud ühte otsa, kuid mitte nii oluliselt kui akrotsentrilistes kromosoomides.
Iga otsadõla kromosoom nimetatakse telomeerideks. Leiti, et telomeeridel on oluline roll kromosoomide stabiilsuse säilitamisel. Telomeerid sisaldavad suurel hulgal TTAGGG nukleotiidjärjestuse kordusi, nn tandemkordusi. Tavaliselt rakkude jagunemise ajal nende korduste arv väheneb telomeerid .
Kuid iga kord valmivad need spetsiaalse ensüümi nntelomeraas... Selle ensüümi aktiivsuse vähenemine viib telomeeride lühenemiseni, mis arvatakse olevat rakusurma põhjus ja mis tavaliselt kaasneb vananemisega.
№77. Denveri kromosoomide klassifikatsioon. Inimese ühtlaselt värvunud kromosoomide klassifikatsioon ja nomenklatuur võeti esmakordselt vastu rahvusvahelisel kohtumisel 1960. aastal Denveris, hiljem mõnevõrra muudeti ja täiendati (London, 1963 ja Chicago, 1966). Denveri klassifikatsiooni järgi jaotatakse kõik inimese kromosoomid 7 rühma, mis on järjestatud nende pikkuse kahanevas järjekorras ja võttes arvesse tsentriooliindeksit (lühikese käe pikkuse ja kogu kromosoomi pikkuse suhe, väljendatuna protsent). Rühmad on tähistatud ingliskeelse tähestiku tähtedega A-st G-ni. Kõik kromosoomipaarid on tavaliselt nummerdatud araabia numbritega. Rühmade omadused on toodud tabelis. 4. Kavandatav klassifikatsioon võimaldas selgelt eristada erinevatesse rühmadesse kuuluvaid kromosoome. Alates 1960. aastast algab kiire kliinilise tsütogeneetika areng: 1959. aastal avastas J. Lejeune Downi sündroomi kromosomaalse olemuse; K. Ford, P. Jacobs ja J. Strong kirjeldasid karüotüübi tunnuseid Klinefelteri ja Turneri sündroomides; 70ndate alguses. avastati Edwardsi ja Patau sündroomide kromosomaalne olemus, "kassi nutu" sündroom; kirjeldas kromosomaalset ebastabiilsust mitmete pärilike sündroomide ja pahaloomuliste haiguste korral. Samal ajal osutus ühtlase värvusega kromosoomide saamise meetodi rakendamine kromosoomide tuvastamisel ebapiisavalt tõhusaks. Denveri klassifikatsiooni puuduseks on see, et homoloogsete paaride eristamine kromosoomide rühmas on sageli ületamatute raskustega.
Tabel 4^ Kromosoomide rühmad ja nende omadused
|
Grupp |
Kromosoomide arv |
Tsentromeeri asukoht |
tsentriooli indeks (%) |
Märge |
|
Suurim metatsentriline |
Pikal õlal võib esineda sekundaarne ahenemine |
|||
|
Suurim submetatsentriline | ||||
|
Suur metatsentriline |
20% lühem kui esimene |
|||
|
Suur submetatsentriline | ||||
|
6-12 ja X-kromosoom |
Keskmine submetatsentriline |
9. kuupäeval esineb sageli sekundaarne kitsendus |
||
|
Keskmine akrotsentriline |
Kõigil sekundaarsetel kitsendustel |
|||
|
Väike metatsentriline |
Sekundaarne ahenemine esineb 10% juhtudest. |
|||
|
Väike submetatsentriline | ||||
|
Väikseim metatsentriline | ||||
|
21-22 ja Y-kromosoom |
Väikseim akrotsentriline |
21. ja 22. sekundaarsed kitsendused |
№78 Valkude biosüntees rakus. Valkude süntees on väga keeruline, mitmeastmeline protsess. See toimub spetsiaalsetes organellides - ribosoomides. Rakk sisaldab suurt hulka ribosoome. Näiteks E. coli's on neid umbes 20 000.
Kuidas toimub valkude süntees ribosoomides?
Valgumolekulid on põhiliselt polüpeptiidahelad, mis koosnevad üksikutest aminohapetest. Kuid aminohapped ei ole piisavalt aktiivsed, et iseseisvalt omavahel seostuda. Seetõttu tuleb aminohapped enne omavahelist ühendamist ja valgumolekuli moodustamist aktiveerida. See aktiveerimine toimub spetsiaalsete ensüümide toimel. Lisaks on igal aminohappel oma ensüüm, mis on sellele spetsiaalselt häälestatud.
Selle (nagu paljude protsesside puhul rakus) energiaallikaks on adenosiintrifosfaat (ATP).
Aktiveerimise tulemusena muutub aminohape labiilsemaks ja seondub sama ensüümi toimel t-RNA-ga.
On oluline, et iga aminohape vastaks rangelt spetsiifilisele t-RNA-le. Ta leiab "oma" aminohappe ja kannab selle ribosoomi. Seetõttu nimetati seda RNA-d transpordi-RNA-ks.
Järelikult sisenevad ribosoomi mitmesugused aktiveeritud aminohapped koos nende t-RNA-dega. Ribosoom on justkui konveier valguahela kokkupanemiseks erinevatest sinna sisenevatest aminohapetest.
Selgub, et samaaegselt t-RNA-ga, millel "istub" tema enda aminohape, saab ribosoom "signaali" tuumas sisalduvalt DNA-lt. Vastavalt sellele signaalile sünteesitakse ribosoomis üks või teine valk, üks või teine ensüüm (kuna ensüümid on valgud).
DNA suunav mõju valgusünteesile ei toimu otse, vaid spetsiaalse vahendaja, RNA selle vormi abil, mida nimetatakse messenger- või messenger-RNA-ks (m-RNA või i-RNA).
Messenger RNA sünteesitakse tuumas joodi toimel DNA mõjul, seetõttu peegeldab selle koostis DNA koostist. RNA molekul on nagu DNA vormi vorm.
Sünteesitud m-RNA siseneb ribosoomi ja justkui annab sellele struktuurile edasi plaani – millises järjekorras tuleb ribosoomi sisenevad aktiveeritud aminohapped omavahel kombineerida, et teatud valk sünteesiks. Vastasel juhul kantakse DNA-s kodeeritud geneetiline informatsioon m-RNA-sse ja seejärel valku.
№79 Mutatsioon (lat.mutio - muutus) - püsiv (st selline, mille võivad pärida antud raku või organismi järeltulijad) genotüübi muutus, mis toimub välis- või sisekeskkonna mõjul. Mõiste lõi Hugo de Vries. Mutatsioonide esinemise protsessi nimetatakse mutageneesiks.
Genoomne: - polüploidiseerumine (organismide või rakkude moodustumine, mille genoomi esindab rohkem kui kaks (3n, 4n, 6n jne) kromosoomikomplekti) ja neuploidsus (heteroploidsus) - kromosoomide arvu muutus, mis ei ole haploidse hulga kordne (vt Inge-Vechtomov, 1989). Sõltuvalt kromosoomikomplektide päritolust eristavad polüploidid allopolüploide, millel on erinevatest liikidest hübridisatsiooni teel saadud kromosoomide komplektid, ja autopolüploide, mille oma genoomi kromosoomikomplektide arv on n-kordne.
Kromosomaalsete mutatsioonidega tekivad üksikute kromosoomide struktuuris suured muutused. Sel juhul toimub ühe või mitme kromosoomi geneetilise materjali osa kadumine (deletsioon) või kahekordistumine (dubleerimine), kromosoomi segmentide orientatsiooni muutumine üksikutes kromosoomides (inversioon), samuti kromosoomide ülekandmine. geneetilise materjali osa ühest kromosoomist teise (translokatsioon) (äärmuslik juhtum - tervete kromosoomide liit, nn Robertsoni translokatsioon, mis on üleminekuvariant kromosoomilt genoomsele mutatsioonile).
Geenitasandil on geenide DNA primaarstruktuuri muutused mutatsioonide mõjul vähem olulised kui kromosomaalsete mutatsioonide puhul, kuid geenimutatsioonid on sagedasemad. Geenimutatsioonide, ühe või mitme nukleotiidi asenduste, deletsioonide ja insertsioonide tulemusena toimuvad geeni erinevate osade translokatsioonid, dubleerimised ja inversioonid. Juhul, kui mutatsiooni toimel muutub ainult üks nukleotiid, räägitakse punktmutatsioonidest. Kuna DNA sisaldab ainult kahte tüüpi lämmastiku aluseid - puriine ja pürimidiine, jagatakse kõik alusasendustega punktmutatsioonid kahte klassi: üleminekud (puriini asendamine puriiniga või pürimidiini asendamine pürimidiiniga) ja transversioon (puriini asendamine pürimidiiniga või vastupidi) . Punktmutatsioonidel on neli võimalikku geneetilist tagajärge: 1) koodoni tähenduse säilimine geneetilise koodi degeneratsiooni tõttu (sünonüümne terminatsioon (nonsense mutatsioon). Geneetilises koodis on kolm mõttetut koodonit: merevaik - UAG, ocp - UAA ja opaal - UGA (sellega kooskõlas nimi ja mutatsioonid, mis põhjustavad mõttetute kolmikute moodustumist - näiteks merevaigu mutatsioon), 4) vastupidine. asendus (stoppkoodon sensskoodoniga).
Vastavalt nende mõjule geeniekspressioonile jagunevad mutatsioonid kahte kategooriasse: aluspaari asenduste tüübi mutatsioonid ja kaadri nihke (kaadrinihe) tüübi mutatsioonid. Viimased on nukleotiidide deletsioonid või insertsioonid, mille arv ei ole kolmekordne, mis on seotud geneetilise koodi kolmiku olemusega.
Primaarset mutatsiooni nimetatakse mõnikord otseseks mutatsiooniks ja mutatsiooni, mis taastab geeni algse struktuuri, nimetatakse pöördmutatsiooniks ehk reversiooniks. Mutantse organismi algse fenotüübi juurde tagasipöördumine mutantse geeni funktsiooni taastumise tõttu ei toimu sageli mitte tõelise pöördumise, vaid sama geeni mõne muu osa või isegi mittealleelse geeni mutatsiooni tõttu. Sel juhul nimetatakse korduvat mutatsiooni supressormutatsiooniks. Geneetilised mehhanismid, mille abil mutantset fenotüüpi alla surutakse, on väga mitmekesised.
Neerumutatsioonid (eosed) on püsivad somaatilised mutatsioonid, mis esinevad taimede kasvupunktide rakkudes. Viib kloonide varieerumiseni. Neid säilitatakse vegetatiivsel paljundamisel. Paljud kultivarid on neerumutatsioonid.
№80. DNA pakkimine kromosoomi. Kromosoomid koosnevad kromatiinist – DNA ja valkude segust vahekorras ~ 1:1. Tänu interaktsioonile valkudega on DNA molekulil kompaktne struktuur. Inimese DNA ühe ahela keskmine pikkus (kui see on venitatud) on 4 cm See tähendab, et iga raku tuum sisaldab umbes 1 meetri DNA-d. DNA molekuli pakkimisel kromosoomis on kolm tasandit (vt joonist): 1. nukleosoomid, nukleosoomi ahel 2. solenoidspiraal 3. silmused Iga selle tüübi jaoks tuntud kaksik-DNA spiraal on "keerdunud" ümber 4 paarist koosneva kompleksi. valgumolekulid moodustavad "helme" - nukleosoomi. Nukleosoomid, mis on omavahel ühendatud DNA molekuli osadega, moodustavad niidi, mis omakorda on keerdunud solenoidi kujul (see on ühe olemasoleva mudeli järgi), üks pööre - 6 nukleosoomi. See struktuur moodustab silmuseid järgmisel pakenditasemel. Sellisel kujul esineb DNA raku tuumas raku kasvu, selle komponentide sünteesi, sealhulgas DNA dubleerimise staadiumis. Kuid rakkude erinevatel eluetappidel on kromatiini kondenseerumine erinev. Rakkude kasvu perioodil kromatiin tuumas ei kondenseeru, selle asukoha kompaktsus on kromosoomi erinevate osade puhul erinev. Rakkude pooldumise protsessi toimudes muutub kromatiin aina tihedamaks ja silmused, millest eespool juttu oli, moodustavad üksteisele tihedalt pakitud "rosette" (ka ühe olemasoleva mudeli järgi), 18 silmust rõnga kohta. Selle tulemusena omandab kromatiin kondenseerunud kromosoomide kujul, mis on meile õpikutest nii hästi teada.
№81. Gametogenees.(Gamete ja kreeka geneesist - päritolu) sugurakkude - sugurakkude arengu ja moodustumise protsess (vt Gametes). G. meessugurakke (spermatosoidid, spermatosoidid) nimetatakse spermatogeneesiks, emassugurakke (ootsüüte) - oogeneesiks. Loomadel ja taimedel kulgeb G. erinevalt, olenevalt meioosi kohast nende organismide elutsüklis.
Mitmerakulistel loomadel esineb G. spetsiaalsetes organites – sugunäärmetes ehk sugunäärmetes (munasarjad, munandid ja hermafrodiitsed sugunäärmed) ning koosneb kolmest põhietapist: 1) primaarsete sugurakkude – gametogoonia (spermatogoonia ja oogoonia) paljunemine järjestikuste mitooside seeria 2) nende rakkude kasvu ja küpsemist nimetatakse nüüd gametotsüütideks (spermatotsüütideks ja ootsüütideks), millel on sarnaselt gametogooniaga täielik (enamasti diploidne) kromosoomide komplekt (vt Kromosoomid). Sel ajal toimub G. peamine sündmus loomadel - gametotsüütide jagunemine meioosi teel, mis põhjustab nendes rakkudes kromosoomide arvu vähenemist (poolitamist) ja nende muutumist haploidseteks rakkudeks (vt Haploid) - spermatiidid ja ootiidid ; 3) sperma (või spermatosoidide) ja munarakkude teket; sel juhul on munad kaetud mitmete embrüonaalsete membraanidega ja spermatosoidid omandavad lipukesed, mis tagavad nende liikuvuse. Paljude loomaliikide emasloomadel lõpeb meioos ja munaraku moodustumine pärast seemnerakkude tungimist munaraku tsütoplasmasse, kuid enne sperma ja munaraku tuumade ühinemist.
Taimedes eraldatakse G. meioosist ja saab alguse haploidsetest rakkudest, eostest (kõrgematel taimedel mikroeosed ja megaspoorid). Eostest areneb taime sugupõlvkond - haploidne gametofüüt, mille suguelundites - gametangia (isane - antheridia (vt. Anteridium), emane - arhegonia (vt Archegonia)) mitoosi teel, G. Erandiks on suguseemnelised ja katteseemnetaimed, milles spermatogenees läheb otse idanevasse mikrospoori – õietolmu rakku. Kõigis madalama ja kõrgema eosega taimedes on G. antheridia rakkudes mitmekordne jagunemine, mille tulemusena moodustub suur hulk väikeseid liikuvaid spermatosoide. G. arhegoonias - ühe, kahe või enama munaraku moodustumine. Taimsete ja katteseemnetaimede puhul koosneb isane G. õietolmu raku tuuma jagamisest (mitoosi teel) generatiivseks ja vegetatiivseks ning generatiivse tuuma edasisest jagunemisest (ka mitoosi teel) kaheks spermatosoidiks. See jagunemine toimub idaneva õietolmu torus. Emane G. katteseemnetaimedes on ühe munaraku isoleerimine mitoosi teel kaheksatuumalises embrüokoti sees. G. peamine erinevus loomades ja taimedes: loomadel ühendab see rakkude muundumise diploididest haploidideks ja haploidsete sugurakkude moodustumist; taimedes taandub G. haploidsetest rakkudest sugurakkude moodustumiseni.
№82 Karüotüüp - teatud bioloogilise liigi (liigi karüotüüp), teatud organismi (individuaalne karüotüüp) või rakuliini (kloon) rakkudele omase kromosoomide komplekti tunnuste kogum (arv, suurus, kuju jne). Täieliku kromosoomikomplekti (karüogrammi) visuaalset esitust nimetatakse mõnikord ka kariotüübiks. Kromosoomide välimus muutub rakutsükli jooksul oluliselt: interfaasi ajal paiknevad kromosoomid tuumas, reeglina despiraliseerunud ja raskesti jälgitavad, seetõttu kasutatakse rakke nende jagunemise ühes etapis - mitoosi metafaasis. karüotüüp. Kariotüübi määramise protseduuriks võib kasutada mis tahes jagunevate rakkude populatsiooni. Inimese karüotüübi määramiseks kasutatakse reeglina perifeerse vere lümfotsüüte, mille üleminekut puhkefaasist G0 proliferatsiooni kutsub esile mitogenafütohemaglutiniini lisamine. Kariotüübi määramiseks võib kasutada ka luuüdi rakke või naha fibroblastide primaarset kultuuri. Metafaasi staadiumis rakkude arvu suurendamiseks lisatakse rakukultuurile vahetult enne fikseerimist kolhitsiinilinokadasool, mis blokeerib mikrotuubulite moodustumise, takistades seeläbi kromatiidide lahknemist raku jagunemise poolustele ja mitoosi lõpulejõudmist.
Pärast fikseerimist värvitakse metafaasi kromosoomide preparaadid ja pildistatakse; mikrograafidest moodustatakse nn süstematiseeritud karüotüüp - homoloogsete kromosoomide paaride nummerdatud komplekt, kromosoomide kujutised on orienteeritud vertikaalselt lühikeste kätega ülespoole, nende nummerdamine toimub suuruse kahanevas järjekorras, sugukromosoomide paar asetatakse komplekti lõppu (vt joon. 1).
Ajalooliselt saadi esimesed mitteüksikasjalikud karüotüübid, mis võimaldasid klassifitseerida kromosoomide morfoloogia järgi, Romanovsky-Giemsa värvimisega, kuid kromosoomide struktuuri täiendav täpsustamine kariotüüpides sai võimalikuks diferentsiaalmeetodite tulekuga. kromosoomide värvimine. Meditsiinilise geneetika kõige sagedamini kasutatav tehnika on G-diferentsiaalne kromosoomide värvimine.
Klassikalise karüotüübi või spetsiifiliste spektriomadustega piirkondade põikmärkide komplekside võrdlemine võimaldab tuvastada nii homoloogseid kromosoome kui ka nende üksikuid piirkondi, mis võimaldab üksikasjalikult määrata kromosomaalseid aberratsioone - kromosoomisiseseid ja kromosoomidevahelisi ümberkorraldusi, millega kaasneb rikkumine. kromosoomi fragmentide järjestusest (deletsioonid, dubleerimised, inversioonid, translokatsioonid). Selline analüüs on meditsiinipraktikas suure tähtsusega, võimaldades diagnoosida mitmeid kromosomaalseid haigusi, mis on põhjustatud nii kariotüüpide jämedatest rikkumistest (kromosoomide arvu kõrvalekalded) kui ka kromosomaalse struktuuri rikkumisest või rakuliste kariotüüpide paljususest kehas. (mosaiikism).
№83 Meioos- redutseeriv jagunemine, küpsemise jagunemine, rakkude pooldumise meetod, mille tulemusel toimub kromosoomide arvu vähenemine (vähenemine) poole võrra ja üks diploidne rakk (sisaldab kahte kromosoomikomplekti) kahele, mis järgneb kiiresti ühele pärast. teisest jagunemisest tekib 4 haploidi (sisaldab ühte kromosoomikomplekti).
Bioloogiline väärtus M ... on järjepidevuse säilitamineKarüotüüp ja seda tüüpi organismide paljudes põlvkondades ning tagades võimaluse kromosoomide ja geenide rekombinatsiooniks) seksuaalprotsessi käigus. M. on üks peamisi pärilikkuse mehhanisme
Meioos koosneb kahest järjestikusest jagunemisest, mille vahel on lühike vahefaas.
Profaas I - esimese jaotuse profaas on väga keeruline ja koosneb viiest etapist:
Leptoteen või leptoneem - kromosoomide pakkimine, DNA kondenseerumine kromosoomide moodustumisega õhukeste niitide kujul (kromosoomid on lühenenud).
Sügoteen või zigoneem - toimub konjugatsioon - homoloogsete kromosoomide kombinatsioon kahest ühendatud kromosoomist koosnevate struktuuride moodustumisega, mida nimetatakse tetraadideks või bivalentsideks, ja nende edasine tihendamine.
Pachytene või pachynema - (pikim staadium) - mõnes kohas on homoloogsed kromosoomid tihedalt ühendatud, moodustades chiasmata. Neis on üle minemine- kohtade vahetus homoloogsete kromosoomide vahel.
Diploteen või diploneem - toimub kromosoomide osaline dekondensatsioon, samal ajal kui osa genoomist saab töötada, toimuvad transkriptsiooni (RNA moodustumise), translatsiooni (valgu süntees) protsessid; homoloogsed kromosoomid jäävad üksteisega seotuks. Mõnel loomal omandavad ootsüütide kromosoomid selles meiootilise profaasi staadiumis iseloomuliku vormi lambiharja kromosoomid.
Diakinees - DNA kondenseerub uuesti nii palju kui võimalik, sünteetilised protsessid peatuvad, tuuma kest lahustub; tsentrioolid lahknevad poolustele; homoloogsed kromosoomid jäävad üksteisega seotuks.
Profaasi I lõpupoole migreeruvad tsentrioolid raku poolustele, moodustuvad lõhustumisspindli niidid, tuumamembraan ja tuumad hävivad.
Metafaas I – kahevalentsed kromosoomid reastuvad piki raku ekvaatorit.
Anafaas I – mikrotuubulid tõmbuvad kokku, bivalentsid jagunevad ja kromosoomid lahknevad poolustele. Oluline on märkida, et sügoteeni kromosoomide konjugatsiooni tõttu lahknevad poolustele terved kromosoomid, millest igaüks koosneb kahest kromatiidist, mitte aga eraldi kromatiididest, nagu mitoos.
Telofaas I – kromosoomid despiraliseeritakse ja tekib tuumaümbris.
Meioosi teine jagunemine järgneb kohe esimesele, ilma väljendunud interfaasita: S-periood puudub, kuna enne teist jagunemist DNA replikatsiooni ei toimu.
II faas - toimub kromosoomide kondenseerumine, rakukeskus jaguneb ja selle jagunemisproduktid lahknevad tuuma poolustele, tuumaümbris hävib, moodustub lõhustumisspindel.
Metafaas II - ühevalentsed kromosoomid (igaüks koosneb kahest kromatiidist) asuvad "ekvaatoril" (tuuma "poolustest" võrdsel kaugusel) ühes tasapinnas, moodustades nn metafaasiplaadi.
Anafaas II - ühevalendid jagunevad jakromatiididpoolustele lahknema.
Telofaas II – kromosoomid despiraliseeritakse ja tekib tuumaümbris.
Selle tulemusena ühestdiploidne rakknelihaploidsed rakud... Juhtudel, kui meioos on seotudgametogenees(näiteks mitmerakulistel loomadel) arengu käigusmunarakud meioosi esimene ja teine jagunemine on teravalt ebaühtlased. Tulemuseks on üks haploidne muna ja kolm nn redutseerimisorganid(esimese ja teise jaotuse abortiivsed tuletised).
№84 Rakkude jagunemine - mitoos
Kui jälgiksite rakkude jagunemist mikroskoobi all, näete järgmist. Esiteks muutub südamiku sees olev materjal väga tihedaks. Saate isegi eristada diskreetseid struktuure - kromosoome. Teate, et inimese somaatilistel (mittesoolistel) rakkudel on 23 paari kromosoome – igast vanema paarist üks kromosoom. Kokku on inimese somaatilistes rakkudes 46 kromosoomi. Kõik 46 asetsevad lahtri ekvatoriaaltasandil. Selles etapis on selgelt näha, et igaüks 46 kromosoomist koosneb tegelikult kahest kromatiidist, mis on üksteisega ühendatud nupuga sarnases piirkonnas. Seda ala on nimetatud tsentromeeriks. Lahtri vastasotstes on kaks tihedat laiku (tsentriooli), millest igaüks moodustab jaotusspindli niidid. Iga ahel liitub kromosoomiga tsentromeeris. Iga kromosoomipaari külge on kinnitatud kaks ahelat – üks igast tsentrioolist. Siis näeksime, et tsentrioolid keerutavad nagu mähised lõhuspindli niite, mille tulemusena kromosoomid lahknevad raku poolustele. Seejärel jagatakse lahtri sisu. See on hämmastav vaatepilt.
Sündmused arenevad järgmise stsenaariumi järgi. Tuleb aru saada, et rakkude jagunemiseks tuleb lahendada väga oluline ülesanne - peab toimuma DNA dubleerimine (replikatsioon), sest just DNA-sse on kodeeritud kogu info selle kohta, mis ja kuidas rakus juhtuma peaks. Pidage meeles, et DNA on kaksikheeliks. Väljaspool spiraali asub suhkru-fosfaatkarkass, milles suhkrud on omavahel seotud kovalentsete sidemetega fosforhappejäägi kaudu. Spiraali sees on orgaanilised lämmastikalused, mis on paarikaupa ühendatud vesiniksidemetega. Ühte kromosoomi salvestatud informatsiooni kopeerimine ja selle info muudatusteta uude rakku ülekandmine on väga lihtne: vaja on lahti kerida DNA kaksikheeliks ja lõhkuda vesiniksidemed, mis hoiavad koos lämmastikaluste paare. Sellest hetkest alates ilmuvad kaks üksikut lämmastikalustega ahelat, mis ulatuvad välja suhkru-fosfaadi selgroost. Pidage meeles, et ühe DNA ahela iga lämmastikku sisaldav alus moodustab alati sidemed teise DNA ahela väga spetsiifilise alusega. Seega seondub adeniin ainult tümiiniga ja guaniin ainult tsütosiiniga. See on lämmastikualuste komplementaarsuse põhimõte. Pidage meeles, et nukleotiidid on molekulid, mis koosnevad orgaanilisest alusest ja fosforüülitud suhkrust. Teoreetiliselt, kui vajalikud nukleotiidid on nukleoplasmas olemas, sobitub iga DNA ahela iga lämmastikualus sobiva paariga. On spetsiifiline ja väga oluline ensüüm DNA polümeraas, mis teeb selle reaktsiooni võimalikuks. Äsja moodustunud tütar-DNA ahela suhkrud on omavahel seotud fosfodiestersidemete kaudu, mille tulemusena moodustub suhkru-fosfaatkarkass - ja nii see juhtuski! Teil on kaks identset kromosoomi, kus teil oli just üks (joonis 2.7). Muidugi, tegelikult on DNA molekuli replikatsiooniprotsess rakus palju keerulisem ja sõltub mitmete ensüümide toimimisest, kuid sellest räägime allpool. raku pooldumine. Esialgu on kromosoomipaarid tuumas despiraliseeritud olekus ja neid ei saa mikroskoobi all eristada. Sellised rakud on väidetavalt interfaasis. Nendes on kromatiinina tuntud geneetiline materjal sassis mass ega ole nii kompaktne kui siis, kui kromosoomid on mikroskoobi all eristatavad. Kromatiin sai oma nime selle järgi, et seda värviti spetsiaalsete värviliste värvainetega tumedaks. See juhtus ammu enne, kui inimesed teadsid selle tegelikust funktsioonist. Nagu juba mainitud, on inimese somaatilistel rakkudel 23 paari kromosoome. Kakskümmend kahte paari nimetatakse autosomaalseteks kromosoomideks, mis on meestel ja naistel samad. Kahekümne kolmandat paari esindavad sugukromosoomid, mis on meestel ja naistel erinevad. Naistel on kaks suurt kromosoomi, mida nimetatakse X-kromosoomideks, samas kui meestel on üks suur X-kromosoom ja üks väike Y-kromosoom. Ühe paari kromosoomidel on sama struktuur. Erinevatelt vanematelt saadud kromosoomid kannavad aga erinevat informatsiooni ja selles mõttes ei ole kromosoomipaarid identsed.
Joonisel fig. 2.8, on universaalne viis eukarüootsete rakkude jagamiseks. Mitoos koosneb neljast peamisest erinevast faasist: profaas, metafaas, anafaas, telofaas. Kui rakk hakkab jagunema, on iga kromosoomi geneetiline materjal juba kahekordistunud. Profaasis vastab kromosoomide arv 46-le, kuid iga kromosoom on kahekordne, see tähendab, et see koosneb kahest sõsarkromatiidist.
Geneetilise materjali kahekordistamise alustamiseks peab rakk lahendama keerulise ülesande – DNA spiraali lahti kerima. Enamik meist arvab, et kromosoom pole nii pikk, sest nii oleme harjunud neid kõigil joonistel nägema. Tegelikkuses on faasidevahelises rakus DNA molekul ülikeerdunud olekus ehk väga tihedalt ja kindlas järjekorras keerdunud. Näib, et kui DNA molekul oleks raku sees keerdumata olekus, siis oleks selle pikkus umbes 1 m! Kujutage vaid ette, kui keeruline ja korrapärane peab olema nii suure hulga geneetilise materjali pakkimine mikroskoopilisse rakku. Sellest saab selgeks, miks eukarüootsetes rakkudes on DNA molekuli kaksikheeliks pakitud kompaktsesse struktuuri, mis moodustub interaktsioonil mitmete valkudega, mida nimetatakse histoonideks. Seega pakivad kromosomaalsed valgud hiiglasliku molekuli vaid mõne kuupmikroni suurusesse tuuma (joonis 2.9) Vahetult enne replikatsiooniprotsessi algust keritakse DNA molekul lahti ensüümi helikaasi toimel. Seejärel katkevad vesiniksidemed, ahelad lahknevad ja nende maatriksile tekivad komplementaarsuse põhimõttel tütarahelad.Rakkude jagunemine on rangelt kontrollitud. On rakke, mis jagunevad sageli. Nende hulka kuuluvad luuüdi rakud. Teised rakud, näiteks luurakud, jagunevad peamiselt elundi aktiivse kasvu või kahjustuse ajal. Lõpuks on rakke, mis jagunevad väga harva või üldse mitte. See tüüp hõlmab küpseid närvirakke. Kontrollimatu rakkude jagunemine ja kasv on märk nende vähist degeneratsioonist. Rakkude jagunemise protsess nõuab regulaatoreid, mida nimetatakse aktiveerimisfaktoriteks. Kasvutegurid on selliste tehaste näited. Kasvufaktorite uurimine on kaasaegse bioloogia ja meditsiini ristumiskohas üks kiiremini kasvavaid valdkondi. Paljud tegurid on praegu esile tõstetud ja osaliselt iseloomustatud.
Aktiveerimisfaktorid töötavad tavaliselt DNA kuju muutmise ja DNA replikatsiooni eest vastutava DNA polümeraasi endaga seondumise kaudu. DNA polümeraas leiab kõigepealt DNA molekulis kindla koha. See silmusekujuline piirkond on vajalik DNA polümeraasi DNA-ga seondumiseks. See omakorda moodustub aktiveerimisfaktori DNA-ga seondumisel. DNA molekuli kohta, kus DNA polümeraas kinnitub ja replikatsiooniprotsess algab, nimetatakse replikatsiooni alguspunktiks. Samuti peaksite teadma, et DNA sünteesi käivitamiseks on vaja lühikesi RNA järjestusi, mis toimivad "praimeritena". Pärast DNA molekuliga seondumist liigub DNA polümeraas mööda emaahelat, viies lõpule tütarahela ehituse vastavalt komplementaarsuse põhimõttele.
DNA polümeraasi üks olulisemaid funktsioone on vigade parandamine äsja sünteesitud DNA ahelas. Valed aluspaarid ära tundes läheb DNA polümeraas ühe sammu tagasi, tütarahelast eemaldatakse vale alus ja sisestatakse õige. Selle tulemusena on DNA replikatsiooni vigade arv ligikaudu üks viga 10-100 miljardi aluspaari kohta. Kuna aga rakud jagunevad piisavalt sageli, kogunevad vead paratamatult. Paljud neist vigadest mõjutavad rakkude elujõulisust. Kui rakud, milles mutatsioon on toimunud, jäävad siiski ellu, hävitab immuunsüsteem need spetsiaalsete selleks kohandatud rakkude abil.
Lubage mul nüüd pöörduda tagasi rakkude jagunemise protsessi juurde. Mitoosi profaasis koosnevad kromosoomid kahest kromatiidist, mis on DNA replikatsiooni tulemus interfaasis. Neid ei ole aga võimalik mikroskoobi all registreerida, kuna profaasis on õdekromatiidid üksteisega tihedas kontaktis, suundudes üksteise suhtes spiraalselt. Prometafaasis tuumamembraan hävib ja kromosoomid hakkavad liikuma raku ekvatoriaaltasandi poole. See protsess tagab raku jagunemisel kromosoomide normaalse eraldumise raku poolustele, mille tulemusena satub igasse vastloodud rakku terviklik kromosoomikomplekt. Ühe kromosoomi sõsarkromatiidid on omavahel seotud punktis, mida nimetatakse tsentromeeriks. Tsentromeeri piirkonnas on kinetokoor, keeruline valgu struktuur, mis on seotud tsentromeeri piirkonna DNA-ga. Sellesse tsooni kinnituvad mitoosi ajal lõhustumisspindli filamendid. Nagu juba täheldasite, paiknevad raku poolustel tsentrioolid, mille arv samuti kahekordistub enne raku jagunemist. Tsentrioolid osalevad lõhustumisspindli filamentide moodustamises. Iga spindli filament kinnitub kinetokoori piirkonnas ühe kromosoomi külge. Mitoosi teises faasis, metafaasis, reastuvad kahest kromatiidist koosnevad kromosoomid raku ekvatoriaaltasandil.
Lõhustumisspindli filamendid tõmbavad kromosoome, mida sellest hetkest nimetatakse tütarkromosoomideks, raku vastaskülgedesse. Faasi, mille jooksul kromosoomid liiguvad erinevates suundades, nimetatakse anafaasiks. Kui kõik läheb plaanipäraselt, jaguneb tsentriool ja õdekromatiidid lahknevad raku poolustele. Kui kromosoomid ei asetse raku ekvatoriaaltasandil õiges järjekorras, võib rakkude jagunemisprotsess katkeda.
Pärast seda, kui kromosoomid on levinud algraku vastasotstesse, jaguneb rakk tsütokineesi tulemusena kaheks. Mitoosi neljandat ja viimast etappi nimetatakse telofaasiks. Nagu juba mainitud, mitoosi teel paljunevaid rakke nimetatakse somaatilisteks. Nende hulka kuuluvad kõik meie keha rakud, välja arvatud rakud, millest hiljem saavad sperma või munarakud. Rakke, mis aitavad kaasa uue organismi moodustumisele, nimetatakse sugurakkudeks. Sugurakud moodustuvad meioosi tulemusena, mida käsitleme järgmises osas.
Mitoos taandub tavaliselt viivitamata. Kuid igal hetkel toimub inimkehas miljoneid rakkude jagunemisi. Aeg-ajalt on rakkudes geneetilist materjali liiga palju, samas kui teistes võib seda olla normist vähem. Tavaliselt need rakud ellu ei jää või immuunsüsteem eemaldab need.
№85 Pärilikud haigused. Pärilike haiguste klassifikatsioon. Pärilikud haigused - haigused, mille esinemine ja areng on seotud sugurakkude kaudu päritud rakkude tarkvara defektidega. Seda terminit kasutatakse seoses polüetoloogiliste haigustega, erinevalt kitsamast rühmast - geneetilistest haigustest.
Sõltuvalt esmase mõjutatud rakkude tüübist eristatakse järgmisi haigusrühmi: sugurakkude mutatsioonidest tingitud haigused - "gametic", s.o. tegelikult pärilikud haigused (näiteks fenüülketonuuria, hemofiilia); need haigused on pärilikud; somaatiliste rakkude mutatsioonidest tingitud haigused - "somaatilised" (näiteks kasvajad, mõned immuunsüsteemi autoagressiooni haigused). Need haigused ei ole päritud; haigused, mis on tingitud idu- ja somaatiliste rakkude mutatsioonide kombinatsioonist (nt perekondlik retinoblastoom).
Eraldi vaadeldakse letaalseid, subletaalseid ja hüpogenitaalseid haigusi: letaalsed haigused põhjustavad emakasisese arengu käigus surma (näiteks autosoomide monosoomia, haploidsus, enamik polüploidsust); subletaalsed haigused põhjustavad inimese surma enne puberteeti (näiteks pärilikud immuunpuudulikkused, nagu Šveitsi tüüpi agam-maglobulineemia, Louis-Bari sündroom, teatud hemofiilia); hüpogenitaalsed haigused kombineeritakse viljatusega (näiteks Shereshevsky-Turneri, Kleinfelteri sündroomid).
Sõltuvalt pärilikkuse ja keskkonna rollist eristatakse nelja haiguste rühma: pärilikud haigused (arenevad mutatsioonide tulemusena); ökogeneetilised haigused (areneb mutatsiooni olemasolul konkreetse faktori keskkonna mõjul); geneetilise eelsoodumusega haigused (geneetiliste tegurite panus on ilmne); keskkonnateguritest põhjustatud haigused (rangelt võttes ei kuulu see haiguste rühm pärilike haiguste hulka, kuna geneetiliste tegurite panus nende esinemisse, arengusse ja tulemustesse võib olla äärmiselt väike, kaheldav või teadmata).