Mendel s-a angajat în selecția mazărelor și tocmai mazărei, norocului științific și rigoarei experimentelor lui Mendel îi datorăm descoperirea legilor de bază ale moștenirii: legea uniformității hibrizilor din prima generație, legea divizării și legea combinarii independente.
Mendel a experimentat cu 22 de soiuri de mazăre, care diferă unele de altele prin 7 caracteristici (culoare, textura semințelor). Mendel și-a condus munca timp de opt ani, a studiat 10.000 de plante de mazăre. Toate formele de mazăre pe care le-a investigat erau de linii pure; rezultatele încrucișării unor astfel de plante între ele au fost întotdeauna aceleași. Mendel a prezentat rezultatele muncii sale într-un articol din 1865, care a devenit piatra de temelie a geneticii. Este greu de spus ce merită mai multă admirație în el și în opera sa - rigoarea experimentelor, claritatea prezentării rezultatelor, cunoașterea perfectă a materialului experimental sau cunoașterea lucrării predecesorilor săi.
Colegii și contemporanii lui Mendel nu au putut să aprecieze importanța concluziilor sale. Potrivit A.E. Gaisinovici, până la sfârșitul secolului al XIX-lea. a fost citat doar de cinci ori și un singur om de știință - botanistul rus I.O. Schmalhausen – a apreciat importanța acestei lucrări. Cu toate acestea, la începutul secolului XX, legile descoperite de el au fost redescoperite aproape simultan și independent unele de altele de oamenii de știință K. Correns, E. Cermak și K. de Vries. Semnificația acestor descoperiri a devenit imediat evidentă pentru comunitatea științifică la începutul anilor 1900; recunoașterea lor a fost asociată cu anumite succese în citologie și formarea ipotezei eredității nucleare.
|
Principalele modele de moștenire a trăsăturilor. Ereditatea umană a fost studiată într-o măsură mult mai mică decât mecanismele de moștenire a trăsăturilor în organismele vegetale și animale. Cu toate acestea, au fost deja obținute date interesante despre dependența ereditară a multor caracteristici fizice ale unei persoane. Nivelul modern al științei genetice ne permite, de asemenea, să afirmăm că toate modelele de bază ale moștenirii trăsăturilor și legile eredității, identificate în experimentele cu plante și animale, sunt valabile și pentru oameni. De exemplu, dacă un tânăr care are gene pentru ochi căprui (homozigot pentru căprui) pe ambii cromozomi omologi se căsătorește cu o fată cu ochi albaștri ai cărei cromozomi conțin, de asemenea, numai gene pentru ochi albaștri, atunci urmașii lor vor moșteni culoarea ochilor conform legilor descoperite. de către fondatorul geneticii G. Mendel încă din anii 60 ai secolului trecut în experimentele sale cu hibrizi de plante de mazăre. Moștenirea sexuală la oameni și la majoritatea animalelor are loc, de asemenea, conform legilor generale și este asociată cu distribuția cromozomilor care au o formă în formă de X și K (cromozomi sexuali). Setul de cromozomi al corpului feminin conține doi cromozomi X, iar corpul masculin conține un cromozom X și un cromozom Y (vezi Fig. 7). Numărul de astfel de exemple poate fi infinit de mare și nu există nicio îndoială că „portretul genetic” al unei persoane (genotip) determină în mare măsură multe dintre proprietățile sale externe (fenotip). Mai jos sunt enumerate câteva dintre trăsăturile ereditare ale unei persoane, determinate de genele dominante și recesive (conform K-Willie, 1974). Manifestarea influenței genelor poate fi efectuată în diferite etape ale ontogenezei, dar majoritatea trăsăturilor fenotipice sunt determinate chiar înainte de naștere. În cele din urmă, însăși manifestarea genelor nu este fatală, ci depinde de factorii de mediu. De exemplu, o boală ereditară severă, fenilcetonuria, nu se dezvoltă dacă un copil care conține genotipurile care provoacă această boală în genotip începe să primească o anumită dietă imediat după naștere. Astfel, genetica are fapte convingătoare care demonstrează existența atât a trăsăturilor strict ereditare (de exemplu, niveluri sanguine, hemofilie, culoarea părului și a ochilor, trăsături faciale și multe altele), cât și a trăsăturilor care sunt în mare măsură determinate de mediul extern (de exemplu, creșterea și masa unei persoane, puterea și dexteritatea mușchilor acestuia, tendința la boală etc.). Trebuie remarcat faptul că nu există o relație directă între gene și trăsături: dezvoltarea unei trăsături poate depinde de influența mai multor gene, iar o genă poate influența dezvoltarea mai multor trăsături. Această imagine este și mai complicată de corecția constantă din mediul extern. În consecință, proprietatea și dezvoltarea populației sunt întotdeauna strâns legate între ele. Este imposibil să ne imaginăm dezvoltarea unui copil fără influențele corective ale mediului, la fel cum este imposibil fără un program de dezvoltare predeterminat genetic, care este o sinteză a experienței istorice a tuturor generațiilor anterioare. Să luăm în considerare rolul eredității și al mediului în dezvoltarea unor caracteristici fizice ale unei persoane în procesul dezvoltării sale prenatale și postnatale. Dezvoltarea prenatală. Formarea organelor și a sistemelor funcționale ale copilului în procesul de embriogeneză este sub controlul genotipului, dar factorii de mediu joacă un rol important. Pentru embrion, organismul matern este mediul primar. Aici, într-un organ special - uterul - embrionul este relativ bine protejat de multe influențe nocive și primește tot ce are nevoie pentru existența sa prin placentă. Cu toate acestea, mai ales în stadiile inițiale de dezvoltare, mulți factori care afectează organismul matern afectează și dezvoltarea embrionului (cele mai semnificative sunt: radiațiile ionizante, bolile suferite de o femeie în timpul sarcinii și multe substanțe chimice: alcool, nicotină, antibiotice, medicamente hormonale etc.). Trebuie remarcat faptul că în perioada prenatală a unei persoane există perioade critice în care organismul în curs de dezvoltare este deosebit de sensibil la acțiunea factorilor externi. Există două astfel de perioade. Prima include debutul dezvoltării prenatale, primele trei săptămâni. În acest moment, are loc depunerea tuturor celor mai importante organe, iar efectele adverse din această perioadă duc adesea la moartea embrionului. În a doua perioadă critică (din a 4-a până în a 7-a săptămână), are loc dezvoltarea ulterioară a tuturor organelor, iar influențele externe dăunătoare în această perioadă pot duce la nașterea unui copil cu diferite defecte fizice (Fig. 9). Este important ca aceleași defecte fizice de naștere să poată fi asociate cu deteriorarea structurilor genetice și cu acțiunea factorilor externi nefavorabili în procesul de embriogeneză. Aceasta este o bună dovadă că mediul și ereditatea sunt în egală măsură responsabile pentru dezvoltarea normală a copilului. Deci, până la momentul nașterii, toate organele unei persoane și sistemele sale fiziologice, inclusiv sistemul nervos, sunt în general formate. În mod firesc, se pune întrebarea: sunt proprietățile sistemului nervos care stau la baza activității mentale a unei persoane, baza minții sale, sunt predeterminate ereditar? Există „preparate pentru conștiință” morfologice gata făcute în același mod în care există pentru activitatea nervoasă inferioară asociată cu reglarea proceselor de viață ale corpului nostru? Pentru a răspunde la aceste întrebări, să ne oprim asupra următoarei etape postnatale a dezvoltării umane. Dezvoltarea postnatală a copilului. În primul rând, vom lua în considerare principalele metode care ne permit să studiem importanța specifică a mediului și a eredității în dezvoltarea postnatală a copiilor și adolescenților. Efectuarea unor astfel de experimente pe plante este simplă și constă în izolarea a două grupe de organisme, identice ca genotip, și plasarea acestor grupuri în condiții diferite de existență. De exemplu, un grup de plante (martor) este cultivat în condiții normale, altul (experimental) - într-o cameră întunecată. În urma unor astfel de experimente, se poate concluziona că formarea clorofilei verzi în plante depinde nu numai de ereditate, ci și de factorii de mediu (lumină), deoarece plantele crescute în întuneric nu vor conține acest pigment. Efectuarea unor experimente similare pe oameni este imposibilă atât din punct de vedere moral și etic, cât și din punct de vedere biologic. Nu există doi oameni în lume cu aceleași genotipuri. Dar... există excepții de la regulă. Aceștia sunt gemeni identici sau homozigoți (IB), având nu numai asemănări externe izbitoare, ci și „portrete” genice aproape identice. Nașterea de gemeni identici și non-identici (TW) nu este neobișnuită; în medie, una din 100 de sarcini se termină cu nașterea a mai mult de un copil. Gemenii este cel mai excelent „material” prezentat omului de știință prin natura însăși, prin urmare metoda în sine este numită geamăn. Care este esența lui? Să presupunem că trebuie să stabiliți care factor este cel mai responsabil pentru dezvoltarea fizică și creșterea unui copil: ereditatea sau mediul? Pentru aceasta se identifică cuplurile de gemeni, se efectuează examinarea morfologică și se studiază stilul de viață (de obicei gemenii, mai ales cei identici, au interese similare și condiții de creștere apropiate). Pe baza studiilor efectuate se stabileste coeficientul de asemanare (concordanta) intre gemeni si se face o concluzie despre rolul ereditatii sau al mediului in dezvoltarea anumitor calitati. Rezultatele unor astfel de studii ale gemenilor identici și neidentici în ceea ce privește frecvența de apariție a unor boli printre aceștia sunt prezentate în tabel. Din păcate, cadrele didactice încă nu lucrează pentru a identifica copiii supradotați, ceea ce este adesea asociat cu o subestimare a individualității fiziologice și mentale a copiilor, cu ignorarea biologiei elementare a unui copil. Munca slabă în identificarea supradotației copiilor poate fi explicată și prin nivelul încă scăzut de cercetare în acest domeniu și lipsa unor criterii științifice precise prin care un profesor ar fi ușor să detecteze înclinațiile ereditare ale unui copil. Există doar o legătură strânsă între supradotație și un nivel ridicat de excitabilitate a sistemului nervos, un simptom al căruia este adesea un dezechilibru ascuțit (psihopatie) al copilului. Profesorul ar trebui să fie deosebit de atent la astfel de copii. Acest lucru este necesar în primul rând din punct de vedere igienic, deoarece acțiunile greșite ale profesorului pot duce la dezvoltarea trăsăturilor de caracter extreme în cele patologice. De asemenea, este necesar pentru identificarea în timp util a înclinațiilor ereditare și dezvoltarea optimă a acestora. Este important să ne amintim că aparenta lipsă de înclinație pentru a învăța, lenea exterioară și indisciplina nu sunt dovezi suficiente ale lipsei de abilități. Există multe dovezi din istorie că această concluzie este adevărată. Creatorul teoriei relativității A. Einstein în copilărie nu a arătat abilități deschise și abia la vârsta de 9 ani a putut să meargă la școala pregătitoare. A. Humboldt (1769-1859) și L. Pasteur (1822-1895), remarcabilul fizician englez I. Newton, erau cunoscuți ca elevi leneși și incapabili la școală. Se poate presupune că tratarea cu pricepere a acestora de către părinți și profesori a creat în cele din urmă toate condițiile necesare dezvoltării talentelor. Mai mult, cea mai importantă calitate a geniului este munca grea. Marele inventator american T. Edison a scris: „Geniul este unul la sută din inspirație, iar nouăzeci și nouă la sută este transpirație”. Cel care nu învață să „transpire” nu va putea niciodată să obțină un mare succes în orice fel de activitate umană, în ciuda celor mai remarcabile înclinații ereditare. Cine, dacă nu un profesor, ar trebui în primul rând să ridice harnicia unui copil - acesta este cel mai important factor în dezvoltarea progresivă a personalității, principala condiție pentru formarea oricăror abilități. Astfel, ereditatea oferă doar „materie primă”, iar mediul își realizează principala „prelucrare”. O persoană care s-a născut chiar și cu cele mai favorabile înclinații, dar trăiește într-un mediu care împiedică dezvoltarea abilităților sale, va rămâne o mediocritate, astfel încât responsabilitatea principală pentru dezvoltarea abilităților intelectuale ale copilului revine educatorilor. Cu toate acestea, rolul mediului nu trebuie absolutizat, nu trebuie uitat că „materialul” furnizat de ereditate nu este întotdeauna de înaltă calitate și nici cel mai talentat sculptor nu va putea sculpta o sculptură din nisip, la fel cum un bloc de marmură de la sine nu va deveni o operă de artă... №67 - nr 68. Moștenire autozomal dominantă- un tip de moștenire în care o alelă mutantă localizată în autozom este suficientă pentru ca boala (sau trăsătura) să fie exprimată. Moștenire autosomal recesivă- tipul de moștenire a unei trăsături sau boli, în care o alelă mutantă localizată într-un autozom trebuie moștenită de la ambii părinți. |
№69. Moștenirea legată de sex - moștenirea unei gene situată în cromozomii sexuali. Moștenirea trăsăturilor care apar numai la indivizii de același sex, dar nu sunt determinate de gene localizate pe cromozomii sexuali, se numește moștenire limitată de sex.
Moștenirea legată de cromozomul X este moștenirea genelor atunci când sexul masculin este heterogametic și se caracterizează prin prezența unui cromozom Y (XY), iar femelele sunt homogametice și au doi cromozomi X (XX). Toate mamiferele (inclusiv oamenii), majoritatea insectelor și reptilelor posedă acest tip de moștenire.
№70 Moștenirea de tip olandez. Holandric heritance (holandricinheritance) - moștenirea unei trăsături controlată de o genă situată într-o parte neomologă a cromozomului Y. Holandricgene este o genă legată de cromozomul Y sexual masculin. Semnele determinate de G. g se transmit numai prin linia masculină - de la tată la fiu.
№71 Cu tip de moștenire legat de X gena mutantă este localizată pe cromozomul X. Dacă mutația are un efect dominant, atunci atât bărbații, cât și femeile pot fi bolnavi. Cu toate acestea, boala se transmite de la un tată bolnav cu o probabilitate de 100% doar la fete, dar nu și la băieți, care primesc un cromozom Y de la tatăl lor (Fig. 5). Probabilitatea transmiterii unei mutații dominante legate de X de la o mamă bolnavă la copii este de 50%. Boala este la fel de probabil să fie moștenită atât de fiică, cât și de fiu.
Pentru dominant,legat de cromozomul X, tipul de moștenire este patologia cunoscută de pediatri, rahitismul rezistent la vitamina D (sinonime: hipofosfatemie, hipofosfatemie familială X-linked, diabet fosfatic). Diagnosticul acestui rahitism sever, care nu dispare sub influența dozelor mari de vitamina D, este confirmat de prezența unei boli similare la unele dintre rude, atât bărbați, cât și femei.
Mai des boli legate de Xsunt moștenite în mod recesiv. O trăsătură distinctivă a bolilor cu recesivăTipul de moștenire legat de X este că bărbații sunt bolnavi în familie și moștenesc alela mutantă de la mama lor practic sănătoasă, heterozigotă pentru alela mutantă. Atunci când alcătuiesc un pedigree, astfel de mame au adesea frați sau unchi bolnavi. Bărbații bolnavi își pot transmite boala doar printr-o generație și numai nepoților (dar nu nepoaților) prin fiica lor sănătoasă, dar heterozigotă.
Astfel, dacă urmăriți în pedigree moștenirea în linia masculinăboala recesiva legata de X, primești ceva de genul „mușcare de cavaler de șah”.
Cel maiboli cunoscute legate de Xsunthemofilia A și B , precum și cea mai gravă patologie a sistemului muscular -Distrofia musculară a lui Duchenne Becker ... Dezvoltarea hemofiliei A se bazează pe mutații ale genei responsabile de sinteza factorului VIII de coagulare a sângelui, iar în hemofilia B, factorul IX de coagulare a sângelui este defect. Ambele gene care codifică factorii VIII și IX sunt localizate pe brațul lung al cromozomului X la q28 și, respectiv, q27.1-2. Se știe că în hemofilie există o încălcare a coagulării sângelui, iar cele mai mici tăieturi pot duce la moarte un pacient fără îngrijiri hematologice speciale. Rețineți că la femei - purtătoare ale genei hemofiliei (așa-numitele „conductori”), în unele cazuri, există și o tendință de sângerare, care se exprimă în menstruație abundentă și sângerare prelungită în timpul nașterii. Această circumstanță trebuie să fie luată în considerare de către obstetricieni-ginecologi atunci când lucrează cu femei care poartă alele mutante în oricare dintre genele hemofiliei A sau B.
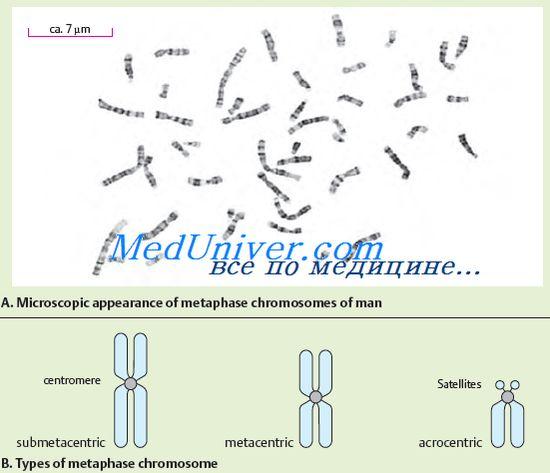
№72 Structura și funcția cromozomilor. Morfologia cromozomului se vede cel mai bine în celulă în stadiul de metafază. Cromozomul este format din două corpuri în formă de baston - cromatide. Ambele cromatide ale fiecărui cromozom sunt identice genetic una cu cealaltă. Cromozomii sunt diferențiați în lungime. Cromozomii au un centromer sau constricție primară, doi telomeri și două brațe. Pe unii cromozomi sunt izolați constricții secundare și sateliți. Mișcarea cromozomului determină Centromerul, care are o structură complexă. ADN-ul centromerului se distinge prin secvența sa de nucleotide caracteristică și proteinele specifice. În funcție de localizarea centromerului, se disting cromozomii acrocentrici, submetacentrici și metacentrici. După cum sa menționat mai sus, unii cromozomi au constricții secundare. Ele, spre deosebire de constricția primară (centromer), nu servesc ca loc de atașare a filamentelor fusului și nu joacă niciun rol în mișcarea cromozomilor. Unele constricții secundare sunt asociate cu formarea nucleolilor, în acest caz se numesc organizatori nucleolari. Organizatorii nucleolari conțin genele responsabile de sinteza ARN. Funcția altor constrângeri secundare nu este încă clară. Unii cromozomi acrocentrici au sateliți – regiuni care sunt conectate cu restul cromozomului printr-un fir subțire de cromatină. Forma și dimensiunea satelitului este constantă pentru un anumit cromozom. La oameni, există sateliți în cinci perechi de cromozomi. Capetele cromozomilor bogati in heterocromatina structurala se numesc telomeri. Telomerii împiedică capetele cromozomilor să se lipească împreună după reduplicare și contribuie astfel la păstrarea integrității lor. În consecință, telomerii sunt responsabili pentru existența cromozomilor ca entități individuale. Cromozomii care au aceeași ordine a genelor se numesc omologi. Au aceeași structură (lungime, localizarea centromerului etc.). Cromozomii neomologi au un set de gene diferit și o structură diferită. Examinarea structurii fine a cromozomilor a arătat că aceștia sunt formați din ADN, proteine și o cantitate mică de ARN. Molecula de ADN poartă sarcini negative distribuite pe toată lungimea sa, iar proteinele atașate de ea - histonele sunt încărcate pozitiv. Acest complex ADN-proteină se numește cromatină. Cromatina poate avea diferite grade de condensare. Cromatina condensată se numește heterocromatina, cromatina decondensată se numește eucromatina. Gradul de decondensare a cromatinei reflectă starea sa funcțională. Regiunile heterocromatinei sunt funcțional mai puțin active decât regiunile eucromatinei, în care majoritatea genelor sunt localizate. Distingeți heterocromatina structurală, a cărei cantitate diferă în cromozomi diferiți, dar este localizată în mod constant în regiunile aproape centromerice. În plus față de heterocromatina structurală, există o heterocromatina opțională care apare în cromozom în timpul supraînfăşurării regiunilor eucromatice. Confirmarea existenței acestui fenomen în cromozomii umani este faptul inactivării genetice a unui cromozom X în celulele somatice ale unei femei. Esența sa constă în faptul că există un mecanism format evolutiv de inactivare a celei de-a doua doze de gene localizate în cromozomul X, drept urmare, în ciuda numărului diferit de cromozomi X în organismele masculine și feminine, numărul de gene. funcționarea în ele este egală. Cromatina este condensată maxim în timpul diviziunii celulare mitotice, apoi poate fi detectată sub formă de cromozomi denși.Dimensiunile moleculelor de ADN ale cromozomilor sunt enorme. Fiecare cromozom este reprezentat de o moleculă de ADN. Pot atinge sute de micrometri și chiar centimetri. Dintre cromozomii umani, cel mai mare este primul; ADN-ul său are o lungime totală de până la 7 cm. Lungimea totală a moleculelor de ADN ale tuturor cromozomilor unei celule umane este de 170 cm. În ciuda dimensiunii gigantice a moleculelor de ADN, este destul de dens împachetat în cromozomi. Această împachetare specifică a ADN-ului cromozomial este asigurată de proteinele histonice. Histonele sunt dispuse pe lungimea moleculei de ADN sub formă de blocuri. Un bloc conține 8 molecule de histonă, formând un nucleozom (o formațiune constând dintr-o catenă de ADN înfășurată în jurul unui octamer de histonă). Nucleozomul are o dimensiune de aproximativ 10 nm. Nucleozomii arată ca niște margele înșirate pe o sfoară. Nucleozomii și secțiunile de ADN care le conectează sunt strâns împachetate sub formă de spirală; există șase nucleozomi pentru fiecare rotație a unei astfel de spirale. Așa se formează structura cromozomului. Informațiile ereditare ale unui organism sunt ordonate strict în funcție de cromozomi individuali. Fiecare organism este caracterizat de un set specific de cromozomi (număr, dimensiune și structură), care se numește cariotip. Cariotipul uman este reprezentat de douăzeci și patru de cromozomi diferiți (22 de perechi de autozomi, cromozomi X și Y). Cariotipul este pașaportul speciei. Analiza cariotipului poate detecta tulburări care pot duce la anomalii de dezvoltare, boli ereditare sau moartea fetușilor și embrionilor în stadiile incipiente de dezvoltare. Multă vreme s-a crezut că cariotipul uman este format din 48 de cromozomi. Cu toate acestea, la începutul anului 1956, a fost publicat un raport, conform căruia numărul de cromozomi din cariotipul uman este de 46. Cromozomii umani diferă în mărime, localizare a centromerilor și constricții secundare. Pentru prima dată, împărțirea cariotipului în grupuri a fost realizată în 1960, la o conferință din Denver (SUA). Descrierea cariotipului uman a inclus inițial următoarele două principii: localizarea cromozomilor pe lungimea lor; gruparea cromozomilor după localizarea centromerului (metacentric, submetacentric, acrocentric). Constanța exactă a numărului de cromozomi, individualitatea lor și complexitatea structurii indică importanța funcției pe care o îndeplinesc. Cromozomii îndeplinesc funcția principalului aparat genetic al celulei. În ele, genele sunt aranjate într-o ordine liniară, fiecare dintre acestea ocupând un loc (locus) strict definit în cromozom. Fiecare cromozom conține multe gene, dar pentru dezvoltarea normală a unui organism este necesar un set de gene dintr-un set complet de cromozomi.
№73. Ereditatea se bazează pe capacitatea tuturor organismelor vii de a acumula, stoca și transmite informații ereditare către urmași. Aceasta una dintre cele mai importante caracteristici calitative ale materiei vii este asociată cu acizii nucleici - acidul dezoxiribonucleic (ADN) și acidul ribonucleic (ARN). Rolul principal îi revine ADN-ului – cea mai lungă moleculă a organismelor vii, concentrată în nucleele celulelor și reprezentând aparatul său ereditar. Lungimea mare a moleculei de ADN face posibilă „scrieți” pe ea, ca pe o bandă telegrafică, toate proprietățile de bază ale viitorului organism și programul de dezvoltare a acestuia. O astfel de „înregistrare” se realizează folosind un „limbaj nucleic” sau „cod nucleic” special, a cărui esență este modificarea secvenței celor patru compuși chimici care alcătuiesc ADN-ul. În mod figurat, „limbajul nucleic” constă din patru litere din care se construiesc cuvinte separate și propoziții întregi ale „limbajului nucleic”.
Pe o astfel de „bandă” nucleică, pot fi distinse zone independente separate, inclusiv o descriere a programului de dezvoltare a unei trăsături. Se numesc gene. Fiecare moleculă de ADN include sute de gene și este un program pentru dezvoltarea multor caracteristici și proprietăți ale organismului. Combinându-se cu proteine speciale și alte substanțe, moleculele de ADN formează formațiuni speciale în nucleu - așa-numiții cromozomi.
Organizarea materialului genetic. Genes [din greacă. genos, naștere] - o unitate de ereditate, o bucată de ADN care ocupă un loc specific în cromozom. Din punctul de vedere al geneticii, o genă este un factor moștenit și o unitate indivizibilă a materialului genetic. Gena structurală (cistron) este un fragment de ADN implicat în formarea unui lanț polipeptidic. Constă dintr-o secvență lider, fragmente de codificare (exoni), secvențe de inserție (nitroni) și o secvență terminală. Deoarece unele proteine constau din mai mult de o subunitate, formularea „o genă – o enzimă” în legătură cu o proteină heteromultimeră (adică formată din două sau mai multe subunități polipeptidice diferite) ar trebui interpretată ca „o genă - un lanț polipeptidic”. .
Genotipul este o colecție de gene dintr-un organism. Chiar și în cele mai vechi timpuri, oamenii foloseau empiric legile moștenirii. Pe baza acestei experiențe, s-a dezvoltat selecția [de la lat. selectio, alege] - știința metodelor de creare a unor noi soiuri de plante și rase de animale prin selecție și încrucișare. Până de curând, genotipul părea inexpugnabil, nesupus acțiunilor umane. Descoperirea structurii genelor a făcut posibilă izolarea lor într-o formă izolată, sintetizarea biochimică și chiar introducerea lor în organism. A devenit posibilă influențarea genei fără a o elimina din organism. Toate acestea au creat premisele pentru manipularea genotipului.
№74 Structura acizilor nucleici.
Acid nucleic(din latină nucleus - nucleus) - un compus organic cu greutate moleculară mare, un biopolimer (polinucleotidă) format din reziduuri de nucleotide. Acizii nucleici ADN și ARN sunt prezenți în celulele tuturor organismelor vii și îndeplinesc cele mai importante funcții pentru stocarea, transmiterea și implementarea informațiilor ereditare. Formele polimerice ale acizilor nucleici se numesc polinucleotide. Lanțurile de nucleotide sunt conectate printr-un rest de acid fosforic (legatură fosfodiester). Deoarece există doar două tipuri de molecule heterociclice în nucleotide, riboză și deoxiriboză, există doar două tipuri de acizi nucleici - acid dezoxiribonucleic (ADN) și acid ribonucleic (ARN).
Formele monomerice se găsesc și în celule și joacă un rol important în semnalizare sau stocarea energiei. Cel mai faimos monomer ARN este ATP, acid adenozin trifosforic, cel mai important acumulator de energie din celulă.
Una dintre funcțiile principale ale acizilor nucleici este de a determina sinteza proteinelor. Informațiile despre structura proteinelor codificate în secvența de nucleotide a ADN-ului trebuie transmise de la o generație la alta și, prin urmare, trebuie copiate corect, adică. sinteza exact aceleiași molecule de ADN (replicare).
№75Replicarea ADN-ului procesul de sinteză a unei molecule fiice de acid dezoxiribonucleic pe matricea moleculei de ADN părinte. În timpul diviziunii ulterioare a celulei mamă, fiecare celulă fiică primește o copie a moleculei de ADN, care este identică cu ADN-ul celulei mamă inițiale. Acest proces asigură transmiterea exactă a informațiilor genetice de la o generație la alta. Replicarea ADN-ului este realizată de un complex enzimatic complex format din 15-20 de proteine diferite, numit repizom. Replicarea ADN-ului este un eveniment cheie în diviziunea celulară. Este esențial ca în momentul divizării ADN-ul să fie replicat complet și o singură dată. Acest lucru este asigurat de anumite mecanisme de reglare a replicării ADN-ului. Replicarea are loc în trei etape:
iniţierea replicării
elongaţie
terminarea replicării.
Reglarea replicării se realizează în principal în etapa de inițiere. Acest lucru este destul de ușor de implementat, deoarece replicarea poate începe nu de la orice bucată de ADN, ci de la una strict definită, numită locul de inițiere a replicării. Pot exista doar unul sau mai multe astfel de site-uri în genom. Conceptul de replicon este strâns legat de conceptul de site de inițiere a replicării. Un replicon este o întindere de ADN care conține un loc de inițiere a replicării și se replic odată ce sinteza ADN-ului începe de la acel loc. Genomul bacteriilor, de regulă, reprezintă un singur replicon, ceea ce înseamnă că replicarea întregului genom este rezultatul unui singur act de inițiere a replicării. Genomul eucariotelor (precum și cromozomii lor individuali) constau dintr-un număr mare de repliconi independenți, ceea ce reduce semnificativ timpul total de replicare al unui cromozom individual. Mecanismele moleculare care controlează numărul de acte de inițiere a replicării la fiecare loc în timpul unui ciclu de diviziune celulară sunt numite control al numărului de copii. Pe lângă ADN-ul cromozomial, celulele bacteriene conțin adesea plasmide, care sunt repliconi individuali. Plasmidele au propriile lor mecanisme de control al copierii: pot asigura sinteza unei singure copii a plasmidei pe ciclu celular sau a mii de copii.
Replicarea începe la locul de inițiere a replicării cu desfășurarea dublei helix ADN, cu formarea unei furculițe de replicare - locul de replicare directă a ADN-ului. Fiecare site poate forma una sau două furcuri de replicare, în funcție de dacă replicarea este unidirecțională sau bidirecțională. Replicarea bidirecțională este mai frecventă. La ceva timp după începerea replicării într-un microscop electronic, este posibil să se observe ochiul de replicare - o secțiune a cromozomului în care ADN-ul a fost deja replicat, înconjurat de secțiuni mai extinse de ADN nereplicat.
În furca de replicare, ADN-ul copiază un complex proteic mare (replicizom), a cărui enzimă cheie este ADN polimeraza. Furculița de replicare se mișcă cu o rată de aproximativ 100.000 de perechi de baze pe minut la procariote și 500-5.000 la eucariote.
№76 Structura și tipurile de cromozomi.Nucleul fiecărei celule somatice a corpului uman conține46 de cromozomi... Kitcromozomii fiecare individ, atât normal cât și patologic, se numește cariotip.
Din46 de cromozomicare alcătuiesc setul de cromozomi umani, 44 sau 22 de perechi reprezintă autozomalcromozomii , ultima pereche sunt cromozomii sexuali. La femei, constituția cromozomilor sexuali este în mod normal reprezentată de doi cromozomi X, iar la bărbați - de cromozomi X și Y.
In toateperechi de cromozomi atât autozomal cât și sexual, unul dintre cromozomi se obține de la tată, iar celălalt de la mamă. Cromozomii unei perechi se numesc omologi sau cromozomi omologi. Celulele de reproducere (sperma și ovule) conțin un set haploid de cromozomi, adică. 23 de cromozomi.
 Spermatozoizii sunt împărțiți în două tipuri, în funcție de conținutcromozomX sau Y. Toate ouăle conțin în mod normalnumai cromozomii
X.
Spermatozoizii sunt împărțiți în două tipuri, în funcție de conținutcromozomX sau Y. Toate ouăle conțin în mod normalnumai cromozomii
X.
Cromozomii sunt clar vizibile după o colorare specială în timpul diviziunii celulare, când cromozomii sunt spiralați maxim. În acest caz, se detectează o constricție în fiecare cromozom, care se numește centromer. Centromerul împarte cromozomul într-un braț scurt (notat cu litera „p”) și un braț lung (notat cu litera „q”). Centromerul determină mișcarea cromozomului în timpul diviziunii celulare. După poziția centromerului, cromozomii sunt clasificați în mai multe grupe. Dacă centromerul este situat în mijlocul cromozomului, atunci un astfel de cromozom se numește metacentric, dacă centromerul este situat mai aproape de unul dintre capetele cromozomului, atunci se numește acrocentric. Unii cromozomi acrocentrici au așa-numiții sateliți, care formează nucleoli într-o celulă nedivizată. Nucleolii conțin numeroase copii ale ARNr. În plus, se disting cromozomii submetacentrici, atunci când centromerul este situat nu în mijlocul cromozomului, ci oarecum deplasat la unul dintre capete, dar nu la fel de semnificativ ca în cromozomii acrocentrici.
Capetele fiecăruiacromozomul umărului numite telomeri. S-a constatat că telomerii joacă un rol important în menținerea stabilității cromozomilor. Telomerii conțin un număr mare de repetări ale secvenței de nucleotide TTAGGG, așa-numitele repetiții în tandem. În mod normal, în timpul diviziunii celulare, există o scădere a numărului acestor repetări în telomerii .
Cu toate acestea, de fiecare dată acestea sunt completate cu ajutorul unei enzime speciale numitetelomeraza... O scădere a activității acestei enzime duce la o scurtare a telomerilor, despre care se crede că este cauza morții celulare, și însoțește în mod normal îmbătrânirea.
№77. Clasificarea cromozomilor Denver. Clasificarea și nomenclatura cromozomilor umani colorați uniform au fost adoptate pentru prima dată la o întâlnire internațională din 1960 la Denver, ulterior oarecum modificate și completate (Londra, 1963 și Chicago, 1966). Conform clasificării Denver, toți cromozomii umani sunt împărțiți în 7 grupe, aranjați în ordinea descrescătoare a lungimii lor și ținând cont de indicele centriol (raportul dintre lungimea brațului scurt și lungimea întregului cromozom, exprimat ca procent). Grupurile sunt desemnate prin litere ale alfabetului englez de la A la G. Toate perechile de cromozomi sunt de obicei numerotate cu cifre arabe. Caracteristicile grupurilor sunt prezentate în tabel. 4. Clasificarea propusă a făcut posibilă distingerea clară între cromozomii aparținând unor grupuri diferite. Din 1960 începe dezvoltarea rapidă a citogeneticii clinice: în 1959, J. Lejeune a descoperit natura cromozomială a sindromului Down; K. Ford, P. Jacobs și J. Strong au descris caracteristicile cariotipului în sindroamele Klinefelter și Turner; la începutul anilor 70. natura cromozomială a sindroamelor Edwards și Patau, a fost descoperit sindromul „plângerii pisicii”; a descris instabilitatea cromozomială într-o serie de sindroame ereditare și boli maligne. În același timp, aplicarea metodei de obținere a cromozomilor uniform colorați s-a dovedit a fi insuficient de eficientă pentru identificarea cromozomilor. Dezavantajul clasificării Denver este că diferențierea perechilor omoloage în cadrul unui grup de cromozomi întâmpină adesea dificultăți insurmontabile.
Tabelul 4^ Grupuri de cromozomi și caracteristicile lor
|
grup |
Numărul cromozomilor |
Locația centromerului |
Indicele centriol (%) |
Notă |
|
Cel mai mare metacentric |
Pe umărul lung, poate exista o constricție secundară |
|||
|
Cel mai mare submetacentric | ||||
|
Metacentric mare |
Cu 20% mai scurt decât primul |
|||
|
Submetacentric mare | ||||
|
6-12 și cromozomul X |
Submetacentric mediu |
Pe 9, apare adesea o constricție secundară |
||
|
Acrocentric mediu |
Pe toate constricțiile secundare |
|||
|
Mic metacentric |
Constricția secundară apare în 10% din cazuri. |
|||
|
Mic submetacentric | ||||
|
Cel mai mic metacentric | ||||
|
21-22 și cromozomul Y |
Cel mai mic acrocentric |
Pe 21 și 22 constricții secundare |
№78 Biosinteza proteinelor în celulă. Sinteza proteinelor este un proces foarte complex, în mai multe etape. Are loc în organele speciale – ribozomi. Celula conține un număr mare de ribozomi. De exemplu, E. coli are aproximativ 20.000 dintre ele.
Cum are loc sinteza proteinelor în ribozomi?
Moleculele de proteine sunt în esență lanțuri polipeptidice compuse din aminoacizi individuali. Dar aminoacizii nu sunt suficient de activi pentru a se lega singuri. Prin urmare, înainte de a se conecta între ele și de a forma o moleculă proteică, aminoacizii trebuie activați. Această activare are loc sub acțiunea unor enzime speciale. În plus, fiecare aminoacid are propria sa enzimă adaptată în mod specific la el.
Sursa de energie pentru aceasta (ca și pentru multe procese din celulă) este adenozin trifosfat (ATP).
Ca urmare a activării, aminoacidul devine mai labil și, sub acțiunea aceleiași enzime, se leagă de t-ARN.
Este important ca fiecare aminoacid să corespundă unui t-ARN strict specific. Ea găsește aminoacidul „ei” și îl transferă în ribozom. Prin urmare, acest ARN a fost numit ARN de transport.
În consecință, diverși aminoacizi activați, combinați cu t-ARN-urile lor, intră în ribozom. Ribozomul este, parcă, o bandă transportoare pentru asamblarea unui lanț proteic din diverși aminoacizi care intră în el.
Se dovedește că simultan cu t-ARN-ul, pe care „se așează propriul aminoacid”, ribozomul primește un „semnal” de la ADN-ul conținut în nucleu. În conformitate cu acest semnal, una sau alta proteină, una sau alta enzimă este sintetizată în ribozom (deoarece enzimele sunt proteine).
Influența direcțională a ADN-ului asupra sintezei proteinelor nu se realizează direct, ci cu ajutorul unui mediator special, acea formă de ARN, care se numește mesager sau ARN mesager (m-ARN sau i-ARN).
ARN-ul mesager este sintetizat în nucleu de iod prin influența ADN-ului, prin urmare compoziția sa reflectă compoziția ADN-ului. Molecula de ARN este ca o matriță sub formă de ADN.
ARNm-ul sintetizat intră în ribozom și, parcă, transmite acestei structuri un plan - în ce ordine aminoacizii activați care intră în ribozom trebuie combinați între ei pentru ca o anumită proteină să fie sintetizată. În caz contrar, informația genetică codificată în ADN este transferată către m-ARN și apoi către proteină.
№79 Mutație (lat.mutatie - schimbare) - modificare persistentă (adică una care poate fi moștenită de descendenții unei anumite celule sau organism) a genotipului care are loc sub influența mediului extern sau intern. Termen inventat de Hugo de Vries. Procesul de apariție a mutațiilor se numește mutageneză.
Genomic: - poliploidizare (formarea de organisme sau celule, al căror genom este reprezentat de mai mult de două (3n, 4n, 6n etc.) seturi de cromozomi) și neuploidie (heteroploidie) - o modificare a numărului de cromozomi care nu este un multiplu al setului haploid (vezi Inge-Vechtomov, 1989). În funcție de originea setului de cromozomi, poliploizii disting între alopoliploizi, care au seturi de cromozomi obținute prin hibridizare de la diferite specii, și autopoliploizi, în care există o creștere a numărului de seturi de cromozomi ai propriului genom, un multiplu de n. .
Cu mutațiile cromozomiale, apar modificări mari în structura cromozomilor individuali. În acest caz, există o pierdere (ștergere) sau dublare a unei părți (duplicare) a materialului genetic al unuia sau mai multor cromozomi, o schimbare a orientării segmentelor cromozomilor în cromozomi individuali (inversie), precum și transferul de o parte a materialului genetic de la un cromozom la altul (translocare) (un caz extrem - unirea cromozomilor întregi, așa-numita translocare Robertsoniană, care este o opțiune de tranziție de la mutația cromozomială la cea genomică).
La nivel de genă, modificările în structura primară a ADN-ului genelor sub influența mutațiilor sunt mai puțin semnificative decât în cazul mutațiilor cromozomiale, cu toate acestea, mutațiile genelor sunt mai frecvente. Ca rezultat al mutațiilor genelor, au loc substituții, deleții și inserții ale uneia sau mai multor nucleotide, translocări, duplicări și inversiuni ale diferitelor părți ale genei. În cazul în care doar o singură nucleotidă se modifică sub acțiunea unei mutații, se vorbește de mutații punctuale. Deoarece ADN-ul conține doar două tipuri de baze azotate - purine și pirimidine, toate mutațiile punctuale cu substituții de baze sunt împărțite în două clase: tranziții (înlocuirea purinei cu purină sau pirimidină cu pirimidină) și transversia (înlocuirea purinei cu pirimidină sau invers) . Există patru consecințe genetice posibile ale mutațiilor punctuale: 1) păstrarea semnificației codonului din cauza degenerării codului genetic (terminare sinonimă (mutație nonsens). Există trei codoni fără sens în codul genetic: chihlimbar - UAG, ocp - UAA și opal - UGA (în conformitate cu aceasta, numele și mutațiile care duc la formarea de tripleți fără sens - de exemplu, mutația chihlimbar), 4) invers înlocuire (codon stop cu codon sens).
În funcție de influența lor asupra expresiei genelor, mutațiile sunt împărțite în două categorii: mutații de tipul substituțiilor de perechi de baze și tipul de schimbare a cadrului (frameshift). Acestea din urmă sunt ștergeri sau inserții de nucleotide, al căror număr nu este un multiplu de trei, ceea ce este asociat cu natura tripletă a codului genetic.
O mutație primară este uneori numită mutație directă, iar o mutație care restabilește structura originală a unei gene se numește mutație inversă sau reversiunea. Revenirea la fenotipul original al organismului mutant datorită restabilirii funcției genei mutante are loc adesea nu datorită unei reversiuni adevărate, ci datorită unei mutații într-o altă parte a aceleiași gene sau chiar a unei alte gene non-alelice. În acest caz, mutația recurentă se numește mutație supresoare. Mecanismele genetice prin care fenotipul mutant este suprimat sunt foarte diverse.
Mutațiile renale (spori) sunt mutații somatice persistente care apar în celulele punctelor de creștere a plantelor. Conduce la variație clonală. Ele se păstrează în timpul înmulțirii vegetative. Multe soiuri sunt mutații renale.
№80. Ambalarea ADN-ului într-un cromozom. Cromozomii sunt compuși din cromatină - un amestec de ADN și proteine într-un raport de ~ 1: 1. Datorită interacțiunii cu proteinele, molecula de ADN are o structură compactă. Lungimea medie a unei catene de ADN uman (dacă este întinsă) este de 4 cm, adică nucleul fiecărei celule conține aproximativ 1 metru de ADN. Ambalarea unei molecule de ADN într-un cromozom are trei niveluri (vezi figura): 1.nucleozomi, o catenă de nucleozom 2. spirală solenoidală 3. bucle O spirală dublă de ADN cunoscută de fiecare dintre tipurile sale „încolăcită” în jurul unui complex de 4 perechi a moleculelor proteice formează o „mărge” – nucleozom. Nucleozomii, interconectați prin secțiuni ale moleculei de ADN, alcătuiesc un fir, care, la rândul său, este răsucit sub forma unui solenoid (acesta este conform unuia dintre modelele existente), o tură - 6 nucleozomi. Această structură formează bucle la nivelul următor de ambalare. În această formă, ADN-ul este prezent în nucleul celulei în stadiul creșterii celulare, sinteza componentelor sale, inclusiv duplicarea ADN-ului. Cu toate acestea, în diferite etape ale vieții celulare, condensarea cromatinei este diferită. În timpul perioadei de creștere a celulelor, cromatina din nucleu nu este condensată, compactitatea locației sale diferă pentru diferite părți ale cromozomului. Când are loc procesul de diviziune celulară, cromatina devine din ce în ce mai densă, iar buclele, care au fost menționate mai sus, formează „rozete” strâns împachetate una cu alta (tot după unul dintre modelele existente), câte 18 bucle pe inel. Ca urmare, cromatina ia forma unor cromozomi condensați, atât de bine cunoscuți nouă din manuale.
№81. Gametogeneza.(de la Gamet și greacă geneza - origine) procesul de dezvoltare și formare a celulelor sexuale - gameți (Vezi Gameți). G. gameții masculini (spermatozoizi, spermatozoizi) se numesc spermatogeneză, gameții feminini (ovocite) - oogeneză. La animale și plante, G. procedează diferit, în funcție de locul meiozei în ciclul de viață al acestor organisme.
La animalele pluricelulare, G. apare în organe speciale - gonade, sau gonade (ovare, testicule și gonade hermafrodite) și constă din trei etape principale: 1) reproducerea celulelor germinale primare - gametogonia (spermatogonie și oogonie) printr-un serie de mitoze succesive 2) creșterea și maturarea acestor celule se numesc acum gametocite (spermatocite și ovocite), care, ca și gametogonia, au un set complet (în mare parte diploid) de cromozomi (vezi Cromozomi). În acest moment, are loc principalul eveniment al G. la animale - divizarea gametocitelor prin meioză, ducând la o reducere (înjumătățire) a numărului de cromozomi din aceste celule și la transformarea lor în celule haploide (vezi Haploide) - spermatide și ootide ; 3) formarea spermatozoizilor (sau spermatozoizilor) și a ovulelor; în acest caz, ouăle sunt îmbrăcate cu o serie de membrane embrionare, iar spermatozoizii dobândesc flageli, care le asigură mobilitatea. La femelele din multe specii de animale, meioza și formarea ovulelor sunt finalizate după pătrunderea spermatozoizilor în citoplasma ovocitului, dar înainte de fuziunea nucleelor spermatozoizilor și a ovulului.
La plante, G. este separat de meioza si incepe in celule haploide, in spori (la plantele superioare, microspori si megaspori). Din spori se dezvoltă generația sexuală a plantei - gametofitul haploid, în ale cărui organe genitale - gametangia (mascul - anteridii (vezi Anteridium), femela - arhegonia (vezi Archegonia)) prin mitoză, G. Excepție fac gimnospermele și angiosperme, în care spermatogeneza merge direct în microsporii germinați - celula de polen. În toate plantele cu spori inferiori și superiori G. în anteridii este o diviziune multiplă a celulelor, în urma căreia se formează un număr mare de spermatozoizi mobili mici. G. în arhegonie - formarea unui, două sau mai multe ovocite. La gimnosperme și angiosperme, masculul G. constă în împărțirea (prin mitoză) a nucleului celulei polenului în generativ și vegetativ și în continuarea divizării (tot prin mitoză) a nucleului generator în doi spermatozoizi. Această diviziune are loc în tubul de polen în germinare. Femela G. la angiosperme este izolarea prin mitoză a unei celule ou în interiorul unui sac embrionar cu opt nuclee. Principala diferență a lui G. la animale și la plante: la animale, combină transformarea celulelor din diploide în haploide și formarea gameților haploizi; la plante, G. se reduce la formarea gametilor din celule haploide.
№82 Cariotip - un set de caracteristici (număr, mărime, formă etc.) ale unui set complet de cromozomi inerente celulelor unei anumite specii biologice (specie cariotip), unui anumit organism (cariotip individual) sau linie (clonă) de celule. Reprezentarea vizuală a unui set complet de cromozomi (cariogramă) este uneori numită și cariotip. Aspectul cromozomilor se modifică semnificativ în timpul ciclului celular: în timpul interfazei, cromozomii sunt localizați în nucleu, de regulă, despiralizați și greu de observat, prin urmare, celulele sunt folosite pentru a determina cariotipul într-una dintre etapele diviziunii lor - metafaza mitozei. Pentru procedura de determinare a cariotipului poate fi utilizată orice populație de celule în diviziune. Pentru a determina cariotipul uman, de regulă, se folosesc limfocite din sângele periferic, a căror tranziție de la stadiul de repaus G0 la proliferare este provocată de adăugarea de mitogenafitohemaglutinină. Celulele măduvei osoase sau cultura primară de fibroblaste cutanate pot fi, de asemenea, utilizate pentru a determina cariotipul. Pentru a crește numărul de celule în stadiul de metafază, colchicinilinocadazolul este adăugat în cultura celulară cu puțin timp înainte de fixare, care blochează formarea microtubulilor, împiedicând astfel cromatidele să diverge către polii diviziunii celulare și finalizarea mitozei.
După fixare, preparatele cromozomilor metafazici sunt colorate și fotografiate; din micrografii se formează așa-numitul cariotip sistematic - un set numerotat de perechi de cromozomi omologi, imaginile cromozomilor sunt orientate vertical cu brațele scurte în sus, numerotarea lor se realizează în ordinea descrescătoare a mărimii, o pereche de cromozomi sexuali este plasată la capătul setului (vezi Fig. 1).
Din punct de vedere istoric, primele cariotipuri nedetaliate care au făcut posibilă clasificarea în funcție de morfologia cromozomilor au fost obținute prin colorarea Romanovsky-Giemsa, cu toate acestea, o detaliere suplimentară a structurii cromozomilor în cariotipuri a devenit posibilă odată cu apariția tehnicilor de colorare cromozomală diferențială. Cea mai des folosită tehnică în genetica medicală este colorarea cromozomilor diferenţial G.
Compararea complexelor de mărci transversale în cariotipia clasică sau regiuni cu caracteristici spectrale specifice face posibilă identificarea atât a cromozomilor omologi, cât și a regiunilor lor individuale, ceea ce face posibilă determinarea în detaliu a aberațiilor cromozomiale - rearanjamente intra și intercromozomiale, însoțite de o încălcare. de ordinul fragmentelor de cromozomi (deleții, duplicări, inversiuni, translocații). O astfel de analiză este de mare importanță în practica medicală, permițând diagnosticarea unui număr de boli cromozomiale cauzate atât de încălcări grave ale cariotipurilor (anomalii ale numărului de cromozomi), cât și de o încălcare a structurii cromozomiale sau a multiplicității cariotipurilor celulare din organism. (mozaicism).
№83 Meioza- diviziune de reducere, diviziune de maturare, o metodă de diviziune celulară, în urma căreia are loc o scădere (reducere) a numărului de cromozomi la jumătate și o celulă diploidă (conținând două seturi de cromozomi) după două care se succed rapid unul după altul diviziunile dau naștere la 4 haploide (conținând un set de cromozomi).
Valoarea biologică M ... este de a menține consistențaCariotip iar într-un număr de generaţii de organisme de acest tip şi asigurând posibilitatea recombinării) cromozomilor şi genelor în timpul procesului sexual. M. este unul dintre mecanismele cheie ale eredităţii
Meioza constă din 2 diviziuni consecutive cu o scurtă interfață între ele.
Profaza I - profaza primei diviziuni este foarte complexă și constă din 5 etape:
Leptoten sau leptoneme - împachetarea cromozomilor, condensarea ADN-ului cu formarea de cromozomi sub formă de fire subțiri (cromozomii sunt scurtați).
Zigoten sau zigoneme - apare conjugarea - combinația de cromozomi omologi cu formarea de structuri formate din doi cromozomi conectați, numiți tetrade sau bivalenți, și compactarea ulterioară a acestora.
Pachyten sau pachinema - (cea mai lungă etapă) - în unele locuri, cromozomii omologi sunt strâns legați, formând chiasma. În ele există trecere peste- schimb de situsuri între cromozomi omologi.
Diploten sau diploneme - are loc decondensarea parțială a cromozomilor, în timp ce o parte a genomului poate funcționa, au loc procese de transcripție (formarea ARN), translație (sinteza proteinelor); cromozomii omologi rămân legați între ei. La unele animale, cromozomii din ovocite în acest stadiu al profazei meiotice capătă o formă caracteristică cromozomi perie de lampă.
Diachineza - ADN-ul se condensează din nou cât mai mult posibil, procesele sintetice se opresc, învelișul nuclear se dizolvă; centriolii diverg către poli; cromozomii omologi rămân legați între ei.
Spre sfârșitul profezei I, centriolii migrează către polii celulei, se formează filamentele fusului de fisiune, membrana nucleară și nucleolii sunt distruse.
Metafaza I - cromozomii bivalenți se aliniază de-a lungul ecuatorului celulei.
Anafaza I - microtubulii se contractă, bivalenții se divid și cromozomii diverg către poli. Este important de reținut că, datorită conjugării cromozomilor din zigoten, cromozomi întregi diverg către poli, fiecare constând din două cromatide, și nu cromatide separate, ca în mitoză.
Telofaza I - cromozomii sunt despiralizați și apare o înveliș nuclear.
A doua diviziune a meiozei urmează imediat prima, fără o interfază pronunțată: perioada S este absentă, deoarece nu există replicare ADN-ului înainte de a doua diviziune.
Profaza II - are loc condensarea cromozomilor, centrul celular se divide și produsele diviziunii sale diverg către polii nucleului, învelișul nuclear este distrus, se formează un fus de fisiune.
Metafaza II - cromozomii univalenți (fiecare format din două cromatide) sunt situați la „ecuator” (la o distanță egală de „polii” nucleului) într-un singur plan, formând așa-numita placă de metafază.
Anafaza II - univalenți se divid șicromatidediverge spre poli.
Telofaza II - cromozomii sunt despiralizați și apare o înveliș nuclear.
Ca urmare, de la unulcelula diploidăpatrucelule haploide... În cazurile în care meioza este asociată cugametogeneza(de exemplu, la animalele pluricelulare), în timpul dezvoltăriiovocite prima și a doua diviziune a meiozei sunt puternic inegale. Ca rezultat, un ou haploid și trei așa-numitele corpuri de reducere(derivate abortive ale primei și a doua diviziuni).
№84 Diviziunea celulară – mitoză
Dacă ați observa diviziunea celulară la microscop, veți vedea următoarele. În primul rând, materialul din interiorul miezului devine foarte dens. Puteți chiar să distingeți între structurile discrete - cromozomi. Știți că celulele somatice umane (non-sex) au 23 de perechi de cromozomi - câte un cromozom din fiecare pereche de la fiecare părinte. În total, există 46 de cromozomi în celulele somatice umane. Toate cele 46 se aliniază în planul ecuatorial al celulei. În această etapă, este clar că fiecare dintre cei 46 de cromozomi constă de fapt din două cromatide, conectate între ele într-o regiune asemănătoare unui buton. Această zonă a fost numită centromer. La capetele opuse ale celulei, există două pete dense (centriole), fiecare dintre acestea formând firele fusului de diviziune. Fiecare catenă se unește cu cromozomul la centromer. La fiecare pereche de cromozomi sunt atașate două fire - câte una de la fiecare centriol. Apoi am vedea că centriolii, asemenea bobinelor, înfășoară firele fusului de fisiune, drept urmare cromozomii diverg către polii celulei. Apoi, conținutul celulei este împărțit. Aceasta este o priveliște uimitoare.
Evenimentele se dezvoltă conform următorului scenariu. Este necesar să înțelegem că pentru diviziunea celulară trebuie rezolvată o sarcină foarte importantă - trebuie să aibă loc duplicarea (replicarea) ADN-ului, deoarece în ADN sunt codificate toate informațiile despre ce și cum ar trebui să se întâmple în celulă. Amintiți-vă că ADN-ul este un dublu helix. În afara spiralei se află coloana vertebrală zahăr-fosfat, în care zaharurile sunt legate între ele prin legături covalente prin reziduul de acid fosforic. În interiorul spiralei există baze organice azotate legate în perechi prin legături de hidrogen. Semnificația copierii informațiilor stocate într-un cromozom și a transferului acestor informații într-o nouă celulă fără modificări este foarte simplă: este necesar să se desfășoare dubla helix ADN și să rupă legăturile de hidrogen care țin împreună perechile de baze azotate. Din acest moment apar două lanțuri simple cu baze azotate, proeminente din coloana vertebrală zahăr-fosfat. Amintiți-vă că fiecare bază azotată a uneia dintre catenele de ADN formează întotdeauna legături cu o bază foarte specifică a celeilalte catene de ADN. Deci, adenina se leagă numai cu timină, iar guanina numai cu citozina. Acesta este principiul complementarității bazelor azotate. De asemenea, rețineți că nucleotidele sunt molecule compuse dintr-o bază organică și zahăr fosforilat. În teorie, dacă nucleotidele necesare sunt prezente în nucleoplasmă, fiecare bază de azot a fiecărei catene de ADN se va potrivi cu o pereche adecvată. Există o enzimă specifică și foarte importantă, ADN polimeraza, care face posibilă această reacție. Zaharurile catenei ADN fiice nou formate sunt conectate între ele prin legături fosfodiesterice, în urma cărora se formează o coloană vertebrală zahăr-fosfat - și așa s-a întâmplat! Aveți doi cromozomi identici unde tocmai ați avut unul (Figura 2.7). Desigur, de fapt, procesul de replicare a unei molecule de ADN într-o celulă este mult mai complicat și depinde de funcționarea unui număr de enzime, dar despre asta vom vorbi mai jos Acum să aruncăm o privire mai atentă asupra procesului de diviziune celulara. Inițial, perechile de cromozomi sunt într-o stare despiralizată în nucleu și nu se pot distinge la microscop. Se spune că astfel de celule sunt în interfază. În acestea, materialul genetic cunoscut sub numele de cromatina este o masă încâlcită și nu este la fel de compact ca atunci când cromozomii se disting la microscop. Cromatina și-a primit numele de la faptul că a fost colorată întuneric cu coloranți colorați speciali. Acest lucru s-a întâmplat cu mult înainte ca oamenii să știe despre adevărata sa funcție. După cum sa menționat deja, celulele somatice umane au 23 de perechi de cromozomi. Douăzeci și două de perechi se numesc cromozomi autozomali, care sunt la fel la bărbați și femele. Cea de-a douăzeci și treia pereche este reprezentată de cromozomi sexuali, care sunt diferiți la bărbați și la femei. Femeile au doi cromozomi mari numiți cromozomi X, în timp ce bărbații au un cromozom X mare și un cromozom Y mic. Cromozomii unei perechi au aceeași structură. Cu toate acestea, cromozomii primiți de la părinți diferiți poartă informații diferite și, în acest sens, perechile de cromozomi nu sunt identice.
Mitoza prezentată în fig. 2.8, este o modalitate universală de împărțire a oricăror celule eucariote. Mitoza constă din patru faze principale distincte: profază, metafază, anafază, telofază. Când o celulă începe să se divizeze, materialul genetic al fiecărui cromozom sa dublat deja. În profază, numărul de cromozomi corespunde cu 46, dar fiecare cromozom este dublu, adică este format din două cromatide surori.
Pentru a începe să dubleze materialul genetic, celula trebuie să rezolve o sarcină dificilă - să desfășoare elica ADN. Cei mai mulți dintre noi considerăm că cromozomul nu este atât de lung, pentru că așa suntem obișnuiți să-i vedem în toate desenele. În realitate, într-o celulă de interfază, molecula de ADN se află într-o stare supercoilată, adică este răsucită foarte strâns și într-o anumită ordine. Indicativ este faptul că, dacă molecula de ADN s-ar afla în interiorul celulei într-o stare nerăsucită, atunci lungimea ei ar fi de aproximativ 1 m! Imaginează-ți cât de complex și ordonat trebuie să fie ambalarea atât de mult material genetic în interiorul unei celule microscopice. Din aceasta devine clar de ce în celulele eucariote dublul helix al moleculei de ADN este împachetat într-o structură compactă formată prin interacțiunea cu un număr de proteine numite histone. Astfel, proteinele cromozomiale împachetează molecula gigantică într-un nucleu cu un volum de doar câțiva microni cubi (Fig. 2.9).Imediat înainte de începerea procesului de replicare, molecula de ADN este derulată de enzima helicaza. Apoi legăturile de hidrogen sunt rupte, lanțurile diverg, iar pe matricea lor se formează lanțuri fiice conform principiului complementarității.Diviziunea celulară este strict controlată. Există celule care se divid frecvent. Acestea includ celulele măduvei osoase. Alte celule, cum ar fi celulele osoase, se divid în principal în timpul creșterii sau lezării organelor active. În cele din urmă, există celule care se divid foarte rar sau deloc. Acest tip include celule nervoase mature. Diviziunea și creșterea celulară necontrolată este un semn al degenerării lor canceroase. Procesul de diviziune celulară necesită regulatori numiți factori de activare. Factorii de creștere sunt exemple de astfel de fabrici. Studiul factorilor de creștere este una dintre zonele cu cea mai rapidă creștere la intersecția dintre biologia și medicina modernă. Mulți factori sunt în prezent evidențiați și parțial caracterizați.
Factorii de activare funcționează de obicei prin schimbarea formei ADN-ului și permițând ADN polimerazei, care este responsabilă pentru replicarea ADN-ului, să se lege de aceasta. ADN polimeraza găsește mai întâi un loc specific în molecula de ADN. Această regiune în formă de buclă este necesară pentru ca ADN polimeraza să se lege de ADN. La rândul său, se formează atunci când factorul de activare se leagă de ADN. Locul moleculei de ADN unde este atașată ADN polimeraza și începe procesul de replicare este numit punctul de origine al replicării. De asemenea, trebuie să știți că sunt necesare secvențe scurte de ARN pentru a iniția sinteza ADN, care funcționează ca „amorsări”. Legăndu-se cu o moleculă de ADN, ADN polimeraza se deplasează de-a lungul lanțului matern, completând construcția lanțului fiice conform principiului complementarității.
Una dintre cele mai importante funcții ale ADN polimerazei este de a corecta erorile din catena de ADN nou sintetizată. Recunoscând perechile de baze greșite, ADN polimeraza se întoarce cu un pas înapoi, baza greșită este îndepărtată din catena fiică și cea corectă este inserată. Ca rezultat, numărul de erori în replicarea ADN-ului este de aproximativ o eroare la 10-100 de miliarde de perechi de baze. Cu toate acestea, deoarece celulele se divid destul de frecvent, erorile se acumulează inevitabil. Multe dintre aceste erori afectează viabilitatea celulară. Dacă celulele în care s-a produs mutația supraviețuiesc totuși, sistemul imunitar le distruge cu ajutorul unor celule speciale adaptate în acest scop.
Să revin acum la procesul de diviziune celulară. În profaza mitozei, cromozomii constau din două cromatide, care este rezultatul replicării ADN-ului în interfază. Cu toate acestea, nu este posibil să le înregistrăm la microscop, deoarece în profază cromatidele surori sunt în contact strâns unele cu altele, spiralând reciproc una față de alta. În prometafază, membrana nucleară este distrusă, iar cromozomii încep să se deplaseze spre planul ecuatorial al celulei. Acest proces asigură separarea normală a cromozomilor de polii celulei în timpul diviziunii celulare, drept urmare un set complet de cromozomi intră în fiecare dintre celulele nou formate. Cromatidele surori ale unui cromozom sunt legate între ele într-un punct numit centromer. În zona centromerului, există un cinetocor, o structură proteică complexă asociată cu ADN-ul regiunii centromerice. Filamentele fusului de fisiune sunt atașate de această zonă în timpul mitozei. După cum ați observat deja, centriolii sunt localizați la polii celulei, numărul cărora se dublează și înainte de diviziunea celulară. Centriolii sunt implicați în formarea filamentelor fusului de fisiune. Fiecare filament fus se atașează la un cromozom din regiunea cinetocorului. În a doua fază a mitozei, metafaza, cromozomii, formați din două cromatide, se aliniază în planul ecuatorial al celulei.
Filamentele fusului de fisiune trag cromozomii, numiți din acest moment cromozomi fiice, la capetele opuse ale celulei. Faza în care cromozomii se mișcă în direcții diferite se numește anafază. Dacă totul decurge conform planului, centriolul se împarte, iar cromatidele surori diverg către polii celulei. Dacă cromozomii nu se aliniază în planul ecuatorial al celulei în ordinea corectă, procesul de diviziune celulară poate fi perturbat.
După ce cromozomii s-au răspândit la capetele opuse ale celulei părinte, celula se împarte în două ca urmare a citokinezei. A patra și ultima etapă a mitozei se numește telofază. Celulele care se înmulțesc prin mitoză, așa cum am menționat deja, sunt numite somatice. Acestea includ toate celulele din corpul nostru, cu excepția celulelor care mai târziu devin spermatozoizi sau ovule. Celulele care contribuie la formarea unui nou organism se numesc celule germinale. Celulele sexuale se formează ca urmare a meiozei, pe care o vom lua în considerare în secțiunea următoare.
De obicei, mitoza se rezolvă fără întârziere. Cu toate acestea, în orice moment, în corpul uman au loc milioane de diviziuni celulare. Din când în când, există prea mult material genetic în celule, în timp ce în altele poate fi mai puțin decât normal. De obicei, aceste celule nu supraviețuiesc sau sunt îndepărtate de sistemul imunitar.
№85 Boli ereditare. Clasificarea bolilor ereditare. Boli ereditare - boli, a căror apariție și dezvoltare este asociată cu defecte în software-ul celulelor, moștenite prin gameți. Termenul este folosit în legătură cu bolile polietiologice, spre deosebire de un grup mai restrâns - bolile genetice.
În funcție de tipul de celule afectate primar, se identifică următoarele grupe de boli: boli datorate mutațiilor în celulele germinale - „gametice”, adică. boli de fapt ereditare (de exemplu, fenilcetonurie, hemofilie); aceste boli sunt moștenite; boli datorate mutațiilor în celulele somatice - „somatice” (de exemplu, tumori, unele boli ale autoagresiunii imune). Aceste boli nu sunt moștenite; boli datorate unei combinații de mutații în celulele germinale și somatice (de exemplu, retinoblastom familial).
Bolile letale, subletale și hipogenitale sunt considerate separat: bolile letale duc la moarte în timpul dezvoltării intrauterine (de exemplu, monosomia prin autozomi, haploidie, majoritatea poliploidiei); bolile subletale duc la moartea individului înainte de pubertate (de exemplu, imunodeficiențe ereditare precum agam-maglobulinemia de tip elvețian, sindromul Louis-Bar, unele hemofilii); bolile hipogenitale sunt combinate cu infertilitatea (de exemplu, sindroamele Shereshevsky-Turner, Kleinfelter).
În funcție de rolul eredității și al mediului, se disting patru grupe de boli: boli ereditare propriu-zise (se dezvoltă ca urmare a mutațiilor); boli ecogenetice (se dezvoltă în prezența unei mutații sub influența unui anumit factor de mediu); boli de predispoziție genetică (aportul factorilor genetici este evident); boli cauzate de factori de mediu (strict vorbind, acest grup de boli nu aparține bolilor ereditare, deoarece contribuția factorilor genetici la apariția, dezvoltarea și rezultatul lor poate fi extrem de mică, îndoielnică sau necunoscută).