Si Mendel ay nakikibahagi sa pagpili ng mga gisantes, at ito ay sa mga gisantes, siyentipikong suwerte at ang hirap ng mga eksperimento ni Mendel na utang natin sa pagtuklas ng mga pangunahing batas ng mana: ang batas ng pagkakapareho ng mga unang henerasyong hybrid, ang batas ng paghahati at ang batas ng malayang kumbinasyon.
Nag-eksperimento si Mendel sa 22 na uri ng mga gisantes, na naiiba sa bawat isa sa 7 katangian (kulay, texture ng buto). Pinangunahan ni Mendel ang kanyang trabaho sa loob ng walong taon, pinag-aralan ang 10,000 na mga halaman ng gisantes. Lahat ng anyo ng mga gisantes na kanyang inimbestigahan ay puro linya; ang mga resulta ng pagtawid sa naturang mga halaman sa bawat isa ay palaging pareho. Iniharap ni Mendel ang mga resulta ng kanyang trabaho sa isang artikulo noong 1865, na naging pundasyon ng genetika. Mahirap sabihin kung ano ang nararapat na higit na paghanga sa kanya at sa kanyang trabaho - ang higpit ng mga eksperimento, ang kalinawan ng presentasyon ng mga resulta, perpektong kaalaman sa eksperimentong materyal o kaalaman sa gawain ng kanyang mga nauna.
Hindi nagawang pahalagahan ng mga kasamahan at kapanahon ni Mendel ang kahalagahan ng kanyang mga konklusyon. Ayon kay A.E. Gaisinovich, hanggang sa katapusan ng ika-19 na siglo. ito ay sinipi lamang ng limang beses, at isang siyentipiko lamang - ang Russian botanist na si I.O. Schmalhausen - pinahahalagahan ang kahalagahan ng gawaing ito. Gayunpaman, sa simula ng ika-20 siglo, ang mga batas na natuklasan niya ay muling natuklasan halos sabay-sabay at independiyente sa bawat isa ng mga siyentipiko na sina K. Correns, E. Cermak at K. de Vries. Ang kahalagahan ng mga pagtuklas na ito ay agad na naging maliwanag sa siyentipikong komunidad noong unang bahagi ng 1900s; ang kanilang pagkilala ay nauugnay sa ilang mga tagumpay sa cytology at ang pagbuo ng hypothesis ng nuclear heredity.
|
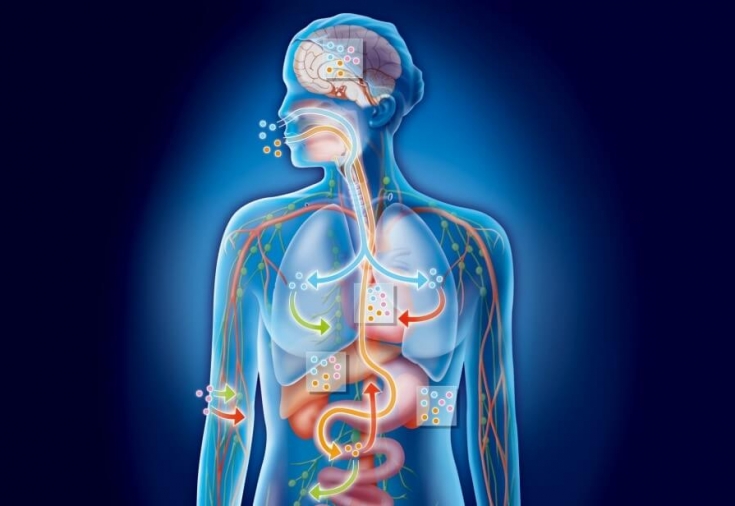
Ang mga pangunahing pattern ng pagmamana ng mga katangian. Ang pagmamana ng tao ay pinag-aralan sa mas maliit na lawak kaysa sa mga mekanismo ng pagmamana ng mga katangian sa mga organismo ng halaman at hayop. Gayunpaman, ang mga kagiliw-giliw na data ay nakuha na sa namamana na pag-asa ng maraming mga pisikal na katangian ng isang tao. Ang modernong antas ng genetic science ay nagpapahintulot din sa amin na igiit na ang lahat ng mga pangunahing pattern ng pagmamana ng mga katangian at ang mga batas ng pagmamana, na tinukoy sa mga eksperimento sa mga halaman at hayop, ay wasto din para sa mga tao. Halimbawa, kung ang isang binata na may mga gene para sa brown na mata (homozygous para sa brown) sa parehong homologous chromosome ay nagpakasal sa isang babaeng may asul na mata na ang mga chromosome ay naglalaman lamang ng mga gene para sa asul na mga mata, kung gayon ang kanilang mga supling ay magmamana ng kulay ng mata ayon sa mga natuklasang batas. ng tagapagtatag ng genetika na si G. Mendel noong 60s ng huling siglo sa kanyang mga eksperimento sa mga hybrid ng halaman ng mga gisantes. Ang pamana ng kasarian sa mga tao at karamihan sa mga hayop ay nangyayari din ayon sa mga pangkalahatang batas at nauugnay sa pamamahagi ng mga chromosome na may hugis-X at K na anyo (sex chromosome). Ang chromosome set ng babaeng katawan ay naglalaman ng dalawang X chromosome, at ang male body ay naglalaman ng isang X chromosome at isang Y chromosome (tingnan ang Fig. 7). Ang bilang ng mga naturang halimbawa ay maaaring walang hanggan na malaki, at walang alinlangan na ang "gene portrait" ng isang tao (genotype) ay higit na tumutukoy sa marami sa kanyang mga panlabas na katangian (phenotype). Nakalista sa ibaba ang ilan sa mga namamana na katangian ng isang tao, na tinutukoy ng dominant at recessive na mga gene (ayon kay K-Willie, 1974). Ang pagpapakita ng impluwensya ng gene ay maaaring isagawa sa iba't ibang yugto ng ontogenesis, ngunit karamihan sa mga phenotypic na katangian ay natutukoy bago pa man ipanganak. Sa wakas, ang mismong pagpapakita ng mga gene ay hindi nakamamatay, ngunit nakasalalay sa mga kadahilanan sa kapaligiran. Halimbawa, ang isang malubhang namamana na sakit, ang phenylketonuria, ay hindi bubuo kung ang isang bata na naglalaman ng mga genotype na sanhi ng sakit na ito sa genotype ay nagsimulang tumanggap ng isang partikular na diyeta kaagad pagkatapos ng kapanganakan. Kaya, ang genetika ay may nakakumbinsi na mga katotohanan na nagpapatunay sa pagkakaroon ng parehong mahigpit na namamana na mga katangian (halimbawa, mga antas ng dugo, hemophilia, kulay ng buhok at mata, mga tampok ng mukha, at marami pang iba), at mga katangiang higit na tinutukoy ng panlabas na kapaligiran (halimbawa, paglaki at masa ng isang tao, ang lakas at kagalingan ng kanyang mga kalamnan, isang pagkahilig sa sakit, atbp.). Dapat pansinin na walang direktang kaugnayan sa pagitan ng mga gene at mga katangian: ang pagbuo ng isang katangian ay maaaring depende sa impluwensya ng maraming mga gene, at ang isang gene ay maaaring makaimpluwensya sa pagbuo ng maraming mga katangian. Ang larawang ito ay mas kumplikado sa pamamagitan ng patuloy na pagwawasto mula sa panlabas na kapaligiran. Dahil dito, ang pagmamay-ari at pag-unlad ng populasyon ay palaging malapit na magkakaugnay. Imposibleng isipin ang pag-unlad ng isang bata nang walang mga corrective na impluwensya ng kapaligiran, tulad ng imposible nang walang genetically predetermined development program, na isang synthesis ng makasaysayang karanasan ng lahat ng nakaraang henerasyon. Isaalang-alang natin ang papel ng pagmamana at kapaligiran sa pagbuo ng ilang pisikal na katangian ng isang tao sa proseso ng kanyang prenatal at postnatal development. Pag-unlad ng prenatal. Ang pagbuo ng mga organo at functional system ng bata sa proseso ng embryogenesis ay nasa ilalim ng kontrol ng genotype, ngunit ang mga kadahilanan sa kapaligiran ay may mahalagang papel. Para sa embryo, ang maternal na organismo ang pangunahing kapaligiran. Dito, sa isang espesyal na organ - ang matris - ang embryo ay medyo protektado mula sa maraming nakakapinsalang impluwensya at natatanggap ang lahat ng kailangan nito para sa pagkakaroon nito sa pamamagitan ng inunan. Gayunpaman, lalo na sa mga unang yugto ng pag-unlad, maraming mga kadahilanan na nakakaapekto sa katawan ng ina ay nakakaapekto rin sa pag-unlad ng embryo (ang pinakamahalaga ay: ionizing radiation, mga sakit na dinaranas ng isang babae sa panahon ng pagbubuntis, at maraming mga kemikal: alkohol, nikotina, antibiotics, hormonal na gamot, atbp.). Dapat pansinin na sa panahon ng prenatal ng isang tao ay may mga kritikal na panahon kapag ang pagbuo ng organismo ay lalong sensitibo sa pagkilos ng mga panlabas na kadahilanan. Mayroong dalawang ganoong panahon. Kasama sa una ang simula ng prenatal development, ang unang tatlong linggo nito. Sa oras na ito, ang pagtula ng lahat ng pinakamahalagang organo ay nangyayari, at ang mga masamang epekto sa panahong ito ay kadalasang humahantong sa pagkamatay ng embryo. Sa ikalawang kritikal na panahon (mula sa ika-4 hanggang ika-7 linggo), ang karagdagang pag-unlad ng lahat ng mga organo ay nangyayari, at ang mga nakakapinsalang panlabas na impluwensya sa panahong ito ay maaaring humantong sa pagsilang ng isang bata na may iba't ibang mga pisikal na depekto (Larawan 9). Mahalaga na ang parehong mga pisikal na depekto sa kapanganakan ay maaaring maiugnay sa pinsala sa mga istrukturang genetic, at sa pagkilos ng hindi kanais-nais na panlabas na mga kadahilanan sa proseso ng embryogenesis. Ito ay magandang katibayan na ang kapaligiran at pagmamana ay pantay na responsable para sa normal na pag-unlad ng bata. Kaya, sa oras ng kapanganakan, ang lahat ng mga organo ng isang tao at ang kanyang mga physiological system, kabilang ang nervous system, ay karaniwang nabuo. Naturally, ang tanong ay lumitaw: ang mga pag-aari ba ng sistema ng nerbiyos na sumasailalim sa aktibidad ng kaisipan ng isang tao, ang batayan ng kanyang pag-iisip, ay namamana na paunang natukoy? Mayroon bang mga handa na morphological "paghahanda para sa kamalayan" sa parehong paraan tulad ng umiiral para sa mas mababang aktibidad ng nerbiyos na nauugnay sa regulasyon ng mga proseso ng buhay ng ating katawan? Upang masagot ang mga tanong na ito, pag-isipan natin ang susunod, postnatal na yugto ng pag-unlad ng tao. Pag-unlad ng bata pagkatapos ng panganganak. Una sa lahat, isasaalang-alang natin ang mga pangunahing pamamaraan na nagpapahintulot sa amin na pag-aralan ang tiyak na kahalagahan ng kapaligiran at pagmamana sa postnatal na pag-unlad ng mga bata at kabataan. Ang pagsasagawa ng gayong mga eksperimento sa mga halaman ay simple at binubuo sa paghihiwalay ng dalawang grupo ng mga organismo, magkapareho sa genotype, at paglalagay ng mga grupong ito sa magkaibang kondisyon ng pag-iral. Halimbawa, ang isang pangkat ng mga halaman (kontrol) ay lumago sa ilalim ng normal na mga kondisyon, isa pa (pang-eksperimento) - sa isang madilim na silid. Bilang isang resulta ng naturang mga eksperimento, maaari itong tapusin na ang pagbuo ng berdeng kloropila sa mga halaman ay nakasalalay hindi lamang sa pagmamana, kundi pati na rin sa mga kadahilanan sa kapaligiran (liwanag), dahil ang mga halaman na lumaki sa dilim ay hindi naglalaman ng pigment na ito. Ang pagsasagawa ng mga katulad na eksperimento sa mga tao ay imposible kapwa mula sa moral at etikal na pananaw, at mula sa biyolohikal. Walang dalawang tao sa mundo na may parehong genotypes. Ngunit ... may mga pagbubukod sa panuntunan. Ang mga ito ay magkapareho, o homozygous, kambal (IB), na hindi lamang may kapansin-pansing panlabas na pagkakatulad, kundi pati na rin ang halos magkaparehong gene na "mga larawan". Ang pagsilang ng identical at non-identical twins (TW) ay hindi karaniwan; sa karaniwan, isa sa 100 pagbubuntis ay nagtatapos sa pagsilang ng higit sa isang bata. Ang Gemini ay ang pinaka mahusay na "materyal" na ipinakita sa siyentipiko sa pamamagitan ng likas na katangian, samakatuwid ang pamamaraan mismo ay tinatawag na kambal. Ano ang kakanyahan nito? Sabihin nating kailangan mong itatag kung aling salik ang pinaka responsable para sa pisikal na pag-unlad at paglaki ng isang bata: pagmamana o kapaligiran? Para dito, natukoy ang mga kambal na mag-asawa, ang kanilang morphological na pagsusuri ay isinasagawa at ang kanilang pamumuhay ay pinag-aaralan (karaniwan ay ang kambal, lalo na ang magkapareho, ay may magkatulad na interes at malapit na kondisyon ng pagpapalaki). Batay sa mga pag-aaral na isinagawa, ang koepisyent ng pagkakatulad (concordance) sa pagitan ng kambal ay itinatag at isang konklusyon ay ginawa tungkol sa papel ng pagmamana o kapaligiran sa pagbuo ng ilang mga katangian. Ang mga resulta ng naturang pag-aaral ng magkapareho at hindi magkatulad na kambal sa mga tuntunin ng dalas ng paglitaw ng ilang mga sakit sa kanila ay ipinakita sa talahanayan. Sa kasamaang palad, ang mga guro ay gumagawa pa rin ng maliit na gawain upang matukoy ang mga bata na may likas na matalino, na kadalasang nauugnay sa isang pagmamaliit sa pisyolohikal at mental na indibidwalidad ng mga bata, na may kamangmangan sa elementarya na biology ng isang bata. Ang mahinang gawain sa pagtukoy sa talento ng mga bata ay maaari ding ipaliwanag ng mababang antas pa rin ng pananaliksik sa lugar na ito at ang kakulangan ng tumpak na pamantayang pang-agham kung saan magiging madali para sa isang guro na tuklasin ang mga namamana na hilig ng isang bata. Mayroon lamang malapit na koneksyon sa pagitan ng pagiging matalino at isang mataas na antas ng excitability ng nervous system, isang sintomas na kung saan ay madalas na isang matalim na kawalan ng timbang (psychopathicity) ng bata. Ang guro ay dapat na maging mas matulungin sa gayong mga bata. Ito ay kinakailangan lalo na mula sa bahagi ng kalinisan, dahil ang mga maling aksyon ng guro ay maaaring humantong sa pag-unlad ng mga matinding katangian ng karakter sa mga pathological. Ito ay kinakailangan din para sa napapanahong pagkakakilanlan ng mga namamana na hilig at ang kanilang pinakamainam na pag-unlad. Mahalagang tandaan na ang maliwanag na kawalan ng hilig na matuto, panlabas na katamaran at kawalan ng disiplina ay hindi sapat na katibayan ng kakulangan ng kakayahan. Maraming ebidensya mula sa kasaysayan na totoo ang konklusyong ito. Ang tagalikha ng teorya ng relativity A. Einstein sa pagkabata ay hindi nagpakita ng mga bukas na kakayahan at sa edad na 9 lamang ay nagawang pumunta sa preparatory school. A. Humboldt (1769-1859) at L. Pasteur (1822-1895), ang namumukod-tanging English physicist na si I. Newton, ay kilala bilang isang tamad at walang kakayahang mag-aaral sa paaralan. Maaari itong ipagpalagay na ang mahusay na paggamot sa kanila ng mga magulang at guro ay nilikha sa dulo ng lahat ng mga kinakailangang kondisyon para sa pag-unlad ng mga talento. Bukod dito, ang pinakamahalagang kalidad ng henyo ay ang pagsusumikap. Ang mahusay na Amerikanong imbentor na si T. Edison ay sumulat: "Ang henyo ay isang porsyento ng inspirasyon, at siyamnapu't siyam na porsyento ay pagpapawis." Ang isang hindi natututong "pawisan" ay hindi kailanman makakamit ang mahusay na tagumpay sa anumang uri ng aktibidad ng tao, sa kabila ng mga pinaka-natitirang namamana na hilig. Sino, kung hindi isang guro, ang dapat una sa lahat ay magdala ng kasipagan sa isang bata - ito ang pinakamahalagang kadahilanan sa progresibong pag-unlad ng pagkatao, ang pangunahing kondisyon para sa pagbuo ng anumang mga kakayahan. Kaya, ang pagmamana ay nagbibigay lamang ng "hilaw na materyal", at ang kapaligiran ay nagsasagawa ng pangunahing "pagproseso" nito. Ang isang tao na ipinanganak kahit na may mga pinaka-kanais-nais na mga hilig, ngunit nakatira sa isang kapaligiran na pumipigil sa pag-unlad ng kanyang mga kakayahan, ay mananatiling isang pangkaraniwan, upang ang pangunahing responsibilidad para sa pag-unlad ng mga intelektwal na kakayahan ng bata ay nahuhulog sa mga tagapagturo. Gayunpaman, ang papel ng kapaligiran ay hindi dapat ganap na ganap, hindi dapat kalimutan na ang "materyal" na ibinibigay ng pagmamana ay hindi palaging may mataas na kalidad, at kahit na ang pinaka-talentadong iskultor ay hindi makakapag-sculpture ng isang iskultura mula sa buhangin, tulad ng isang bloke ng marmol sa kanyang sarili ay hindi magiging isang gawa ng sining ... №67 - hindi. 68. Autosomal na nangingibabaw na mana- isang uri ng pamana kung saan ang isang mutant allele na naka-localize sa autosome ay sapat na para maipahayag ang sakit (o katangian). Autosomal recessive inheritance- ang uri ng pagmamana ng isang katangian o sakit, kung saan ang isang mutant allele na naisalokal sa isang autosome ay dapat na minana mula sa parehong mga magulang. |
№69. Pamana na nauugnay sa kasarian - pagmamana ng isang gene na matatagpuan sa mga sex chromosome. Ang pagmamana ng mga katangian na lumilitaw lamang sa mga indibidwal ng parehong kasarian, ngunit hindi tinutukoy ng mga gene na matatagpuan sa mga chromosome ng kasarian, ay tinatawag na sex-limited inheritance.
Ang inheritance na naka-link sa X chromosome ay ang inheritance ng mga gene kapag ang male sex ay heterogametic at nailalarawan sa pagkakaroon ng Y chromosome (XY), at ang mga babae ay homogametic at may dalawang X chromosome (XX). Lahat ng mammal (kabilang ang mga tao), karamihan sa mga insekto at reptilya ay nagtataglay ng ganitong uri ng pamana.
№70 Hollandric na uri ng mana. Holandric inheritance (holandricinheritance) - pagmamana ng isang katangian na kinokontrol ng isang gene na matatagpuan sa isang hindi homologous na bahagi ng Y-chromosome. Ang holandricgene ay isang gene na naka-link sa male sex Y chromosome. Ang mga palatandaan na tinutukoy ni G. g ay ipinadala lamang sa pamamagitan ng linya ng lalaki - mula sa ama hanggang sa anak na lalaki.
№71 Na may X-linked inheritance type ang mutant gene ay matatagpuan sa X chromosome. Kung ang mutation ay may nangingibabaw na epekto, kung gayon ang mga lalaki at babae ay maaaring magkasakit. Gayunpaman, ang sakit ay nakukuha mula sa isang may sakit na ama na may 100% na posibilidad lamang sa mga batang babae, ngunit hindi sa mga lalaki, na tumatanggap ng Y chromosome mula sa kanilang ama (Larawan 5). Ang posibilidad ng paghahatid ng isang nangingibabaw na X-linked mutation mula sa isang may sakit na ina sa mga bata ay 50%. Ang sakit ay pantay na malamang na namamana ng parehong anak na babae at lalaki.
Sa nangingibabaw,naka-link sa X chromosome, ang uri ng mana ay ang patolohiya na kilala sa mga pediatrician, bitamina D-resistant rickets (mga kasingkahulugan: hypophosphatemia, familial-X-linked hypophosphatemia, phosphatic diabetes). Ang diagnosis ng malubhang rickets na ito, na hindi nawawala sa ilalim ng impluwensya ng malalaking dosis ng bitamina D, ay nakumpirma ng pagkakaroon ng isang katulad na sakit sa ilan sa mga kamag-anak, kapwa lalaki at babae.
Mas madalas na X-linked na mga sakitay minana sa isang recessive na paraan. Isang natatanging katangian ng mga sakit na may recessiveX-linked inheritance type ay ang mga lalaki ay may sakit sa pamilya, at minana nila ang mutant allele mula sa kanilang halos malusog na ina, heterozygous para sa mutant allele. Kapag nag-iipon ng isang pedigree, ang gayong mga ina ay kadalasang may mga kapatid na lalaki o tiyuhin na may sakit. Ang mga lalaking may sakit ay maaaring magpadala ng kanilang sakit sa pamamagitan lamang ng isang henerasyon at sa mga apo lamang (ngunit hindi mga apo) sa pamamagitan ng kanilang malusog ngunit heterozygous na anak na babae.
Kaya, kung i-trace mo sa pedigree inheritance sa male linerecessive X-linked na sakit, nakakakuha ka ng parang "chess knight's move".
Karamihanmga kilalang sakit na nauugnay sa Xayhemophilia A at B , pati na rin ang pinakamalubhang patolohiya ng muscular system -Muscular dystrophy ni Duchenne Becker ... Ang pagbuo ng hemophilia A ay batay sa mga mutasyon sa gene na responsable para sa synthesis ng factor VIII ng blood coagulation, at sa hemophilia B, ang factor IX ng blood clotting ay may depekto. Ang parehong mga gene na nag-encode ng mga kadahilanan VIII at IX ay matatagpuan sa mahabang braso ng X chromosome sa q28 at q27.1-2, ayon sa pagkakabanggit. Ito ay kilala na sa hemophilia mayroong isang paglabag sa pamumuo ng dugo, at ang pinakamaliit na pagbawas ay maaaring humantong sa isang pasyente na walang espesyal na pangangalaga sa hematological hanggang sa kamatayan. Tandaan na sa mga kababaihan - mga carrier ng hemophilia gene (ang tinatawag na "konduktor"), sa ilang mga kaso, mayroon ding posibilidad na dumudugo, na ipinahayag sa mabigat na regla at matagal na pagdurugo sa panahon ng panganganak. Ang sitwasyong ito ay dapat isaalang-alang ng mga obstetrician-gynecologist kapag nagtatrabaho sa mga babaeng carrier ng mutant alleles sa alinman sa mga hemophilia A o B na gene.
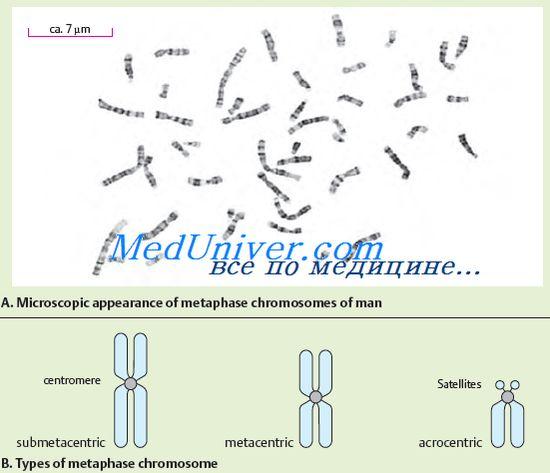
№72 Ang istraktura at paggana ng mga chromosome. Ang chromosome morphology ay pinakamahusay na nakikita sa cell sa yugto ng metaphase. Ang chromosome ay binubuo ng dalawang katawan na hugis baras - chromatids. Parehong chromatid ng bawat chromosome ay genetically identical sa isa't isa. Naiiba ang haba ng mga chromosome. Ang mga chromosome ay may sentromere o pangunahing constriction, dalawang telomere, at dalawang braso. Sa ilang chromosome, ang mga pangalawang constriction at satellite ay nakahiwalay. Tinutukoy ng paggalaw ng chromosome ang Centromere, na may kumplikadong istraktura. Ang sentromere DNA ay nakikilala sa pamamagitan ng katangian nitong pagkakasunud-sunod ng nucleotide at mga tiyak na protina. Depende sa lokasyon ng centromere, ang acrocentric, submetacentric at metacentric chromosome ay nakikilala. Tulad ng nabanggit sa itaas, ang ilang mga chromosome ay may pangalawang constriction. Ang mga ito, sa kaibahan sa pangunahing constriction (centromere), ay hindi nagsisilbing isang lugar ng attachment ng mga spindle filament at hindi gumaganap ng anumang papel sa paggalaw ng mga chromosome. Ang ilang mga pangalawang constriction ay nauugnay sa pagbuo ng nucleoli, sa kasong ito sila ay tinatawag na nucleolar organizers. Ang mga nucleolar organizer ay naglalaman ng mga gene na responsable para sa RNA synthesis. Ang pag-andar ng iba pang mga pangalawang paghihigpit ay hindi pa malinaw. Ang ilang acrocentric chromosome ay may mga satellite — mga rehiyon na konektado sa natitirang bahagi ng chromosome sa pamamagitan ng manipis na thread ng chromatin. Ang hugis at sukat ng satellite ay pare-pareho para sa isang naibigay na chromosome. Sa mga tao, mayroong mga satellite sa limang pares ng chromosome. Ang mga dulo ng chromosome na mayaman sa structural heterochromatin ay tinatawag na telomeres. Pinipigilan ng mga Telomeres ang mga dulo ng chromosome na magkadikit pagkatapos ng reduplication at sa gayon ay nakakatulong sa pangangalaga ng kanilang integridad. Dahil dito, ang mga telomere ay may pananagutan sa pagkakaroon ng mga chromosome bilang mga indibidwal na nilalang. Ang mga kromosom na may parehong pagkakasunud-sunod ng gene ay tinatawag na homologous. Mayroon silang parehong istraktura (haba, lokasyon ng sentromere, atbp.). Ang mga non-homologous chromosome ay may ibang gene set at ibang istraktura. Ang pagsusuri sa pinong istraktura ng mga chromosome ay nagpakita na sila ay binubuo ng DNA, protina at isang maliit na halaga ng RNA. Ang molekula ng DNA ay nagdadala ng mga negatibong singil na ipinamamahagi sa buong haba nito, at ang mga protina na nakakabit dito - ang mga histone ay positibong sinisingil. Ang DNA-protein complex na ito ay tinatawag na chromatin. Maaaring magkaroon ng iba't ibang antas ng condensation ang Chromatin. Ang condensed chromatin ay tinatawag na heterochromatin, ang decondensed chromatin ay tinatawag na euchromatin. Ang antas ng chromatin decondensation ay sumasalamin sa functional state nito. Ang mga rehiyon ng heterochromatin ay hindi gaanong aktibo kaysa sa mga rehiyon ng euchromatin, kung saan ang karamihan sa mga gene ay naisalokal. Makilala ang pagkakaiba sa pagitan ng structural heterochromatin, ang halaga nito ay naiiba sa iba't ibang chromosome, ngunit ito ay patuloy na matatagpuan sa malapit-centromeric na mga rehiyon. Bilang karagdagan sa structural heterochromatin, mayroong isang opsyonal na heterochromatin na lumilitaw sa chromosome sa panahon ng supercoiling ng mga euchromatic na rehiyon. Ang pagkumpirma ng pagkakaroon ng hindi pangkaraniwang bagay na ito sa mga chromosome ng tao ay ang katotohanan ng genetic inactivation ng isang X chromosome sa mga somatic cells ng isang babae. Ang kakanyahan nito ay nakasalalay sa katotohanan na mayroong isang ebolusyonaryong nabuo na mekanismo ng hindi aktibo ng pangalawang dosis ng mga gene na naisalokal sa X chromosome, bilang isang resulta kung saan, sa kabila ng magkaibang bilang ng mga X chromosome sa lalaki at babae na organismo, ang bilang ng mga gene. ang paggana sa kanila ay pantay. Ang Chromatin ay pinakamataas na naka-condensed sa panahon ng mitotic cell division, pagkatapos ay matutukoy ito sa anyo ng mga siksik na chromosome. Ang mga sukat ng mga molekula ng DNA ng mga chromosome ay napakalaki. Ang bawat chromosome ay kinakatawan ng isang molekula ng DNA. Maaari silang umabot ng daan-daang micrometer at kahit na sentimetro. Sa mga chromosome ng tao, ang pinakamalaki ay ang una; ang DNA nito ay may kabuuang haba na hanggang 7 cm. Ang kabuuang haba ng mga molekula ng DNA ng lahat ng chromosome ng isang cell ng tao ay 170 cm. Sa kabila ng napakalaking sukat ng mga molekula ng DNA, ito ay medyo siksik sa mga chromosome. Ang partikular na pag-iimpake ng chromosomal DNA ay ibinibigay ng mga protina ng histone. Ang mga histone ay nakaayos kasama ang haba ng molekula ng DNA sa anyo ng mga bloke. Ang isang bloke ay naglalaman ng 8 histone molecule, na bumubuo ng isang nucleosome (isang pormasyon na binubuo ng isang DNA strand na sugat sa paligid ng isang histone octamer). Ang nucleosome ay halos 10 nm ang laki. Ang mga nucleosome ay mukhang mga kuwintas na nakatali sa isang string. Ang mga nucleosome at ang mga seksyon ng DNA na nag-uugnay sa kanila ay mahigpit na nakaimpake sa anyo ng isang spiral; mayroong anim na nucleosome para sa bawat pagliko ng naturang spiral. Ito ay kung paano nabuo ang istraktura ng chromosome. Ang namamana na impormasyon ng isang organismo ay mahigpit na iniutos ayon sa mga indibidwal na chromosome. Ang bawat organismo ay nailalarawan sa pamamagitan ng isang tiyak na hanay ng mga chromosome (bilang, laki at istraktura), na tinatawag na karyotype. Ang karyotype ng tao ay kinakatawan ng dalawampu't apat na magkakaibang chromosome (22 pares ng autosomes, X at Y chromosomes). Ang karyotype ay ang pasaporte ng mga species. Ang pagsusuri ng karyotype ay maaaring makakita ng mga karamdaman na maaaring humantong sa mga abnormalidad sa pag-unlad, mga namamana na sakit o pagkamatay ng mga fetus at embryo sa mga unang yugto ng pag-unlad. Sa loob ng mahabang panahon, pinaniniwalaan na ang karyotype ng tao ay binubuo ng 48 chromosome. Gayunpaman, sa simula ng 1956, isang ulat ang nai-publish, ayon sa kung saan ang bilang ng mga chromosome sa karyotype ng tao ay 46. Ang mga chromosome ng tao ay naiiba sa laki, lokasyon ng mga sentromere at pangalawang constriction. Sa unang pagkakataon, ang paghahati ng karyotype sa mga grupo ay isinagawa noong 1960 sa isang kumperensya sa Denver (USA). Ang paglalarawan ng karyotype ng tao ay orihinal na kasama ang sumusunod na dalawang prinsipyo: ang lokasyon ng mga chromosome sa haba ng mga ito; pagpapangkat ng mga chromosome ayon sa lokasyon ng centromere (metacentric, submetacentric, acrocentric). Ang eksaktong katatagan ng bilang ng mga chromosome, ang kanilang sariling katangian at ang pagiging kumplikado ng istraktura ay nagpapahiwatig ng kahalagahan ng pag-andar na kanilang ginagawa. Ginagawa ng mga Chromosome ang pag-andar ng pangunahing genetic apparatus ng cell. Sa kanila, ang mga gene ay nakaayos sa isang linear na pagkakasunud-sunod, na ang bawat isa ay sumasakop sa isang mahigpit na tinukoy na lugar (locus) sa chromosome. Ang bawat chromosome ay naglalaman ng maraming mga gene, ngunit para sa normal na pag-unlad ng isang organismo, isang set ng mga gene mula sa isang kumpletong set ng chromosome ay kinakailangan.
№73. Ang pagmamana ay batay sa kakayahan ng lahat ng nabubuhay na organismo na mag-ipon, mag-imbak at magpadala ng namamana na impormasyon sa mga supling. Ang isa sa pinakamahalagang katangian ng husay ng bagay na may buhay ay nauugnay sa mga nucleic acid - deoxyribonucleic acid (DNA) at ribonucleic acid (RNA). Ang nangungunang papel ay kabilang sa DNA - ang pinakamahabang molekula ng mga nabubuhay na organismo, na puro sa nuclei ng mga selula at kumakatawan sa namamana nitong kagamitan. Ang mahabang haba ng molekula ng DNA ay ginagawang posible na "isulat" dito, tulad ng sa isang telegraph tape, ang lahat ng mga pangunahing katangian ng hinaharap na organismo at ang programa ng pag-unlad nito. Ang nasabing "pagre-record" ay isinasagawa gamit ang isang espesyal na "nucleic language", o "nucleic code", na ang kakanyahan nito ay upang baguhin ang pagkakasunud-sunod ng apat na kemikal na compound na bumubuo sa DNA. Sa matalinghagang pagsasalita, ang "nucleic language" ay binubuo ng apat na letra kung saan binuo ang mga hiwalay na salita at mga buong pangungusap ng "nucleic language".
Sa naturang nucleic na "strip", ang hiwalay na mga independiyenteng lugar ay maaaring makilala, kabilang ang isang paglalarawan ng programa ng pag-unlad ng isang katangian. Tinatawag silang mga gene. Ang bawat molekula ng DNA ay may kasamang daan-daang mga gene at isang programa para sa pagbuo ng maraming katangian at katangian ng organismo. Ang pagsasama sa mga espesyal na protina at ilang iba pang mga sangkap, ang mga molekula ng DNA ay bumubuo ng mga espesyal na pormasyon sa nucleus - ang tinatawag na mga chromosome.
Organisasyon ng genetic na materyal. Genes [mula sa Griyego. genos, birth] - isang yunit ng pagmamana, isang piraso ng DNA na sumasakop sa isang tiyak na lugar sa chromosome. Mula sa punto ng view ng genetika, ang isang gene ay isang minanang kadahilanan at isang hindi mahahati na yunit ng genetic na materyal. Ang istrukturang gene (cistron) ay isang fragment ng DNA na kasangkot sa pagbuo ng isang polypeptide chain. Binubuo ito ng isang leader sequence, coding fragment (exons), insertion sequence (nitrons), at isang terminal sequence. Dahil ang ilang mga protina ay binubuo ng higit sa isang subunit, ang mga salitang "isang gene - isang enzyme" na may kaugnayan sa isang heteromultimeric (iyon ay, binubuo ng dalawa o higit pang magkakaibang polypeptide subunits) na protina ay dapat bigyang kahulugan bilang "isang gene - isang polypeptide chain" .
Ang genotype ay isang koleksyon ng mga gene sa isang organismo. Kahit noong sinaunang panahon, empirically ginamit ng mga tao ang mga batas ng mana. Sa batayan ng karanasang ito, nabuo ang pagpili [mula sa lat. selectio, pumili] - ang agham ng mga pamamaraan ng paglikha ng mga bagong uri ng mga halaman at lahi ng hayop sa pamamagitan ng pagpili at pagtawid. Hanggang sa kamakailan lamang, ang genotype ay tila hindi magagapi, hindi napapailalim sa mga aksyon ng tao. Ang pagtuklas ng istraktura ng mga gene ay naging posible upang ihiwalay ang mga ito sa isang nakahiwalay na anyo, synthesize ang mga ito sa biochemically, at kahit na ipakilala ang mga ito sa katawan. Naging posible na maimpluwensyahan ang gene nang hindi inilalabas ito mula sa katawan. Ang lahat ng ito ay lumikha ng mga kinakailangan para sa pagmamanipula ng genotype.
№74 Ang istraktura ng mga nucleic acid.
Nucleic acid(mula sa Latin na nucleus - nucleus) - isang mataas na molekular na timbang na organic compound, isang biopolymer (polynucleotide) na nabuo sa pamamagitan ng mga nalalabi ng nucleotides. Ang mga nucleic acid na DNA at RNA ay naroroon sa mga selula ng lahat ng buhay na organismo at gumaganap ng pinakamahalagang tungkulin para sa pag-iimbak, paghahatid at pagpapatupad ng namamana na impormasyon. Ang mga polymeric na anyo ng mga nucleic acid ay tinatawag na polynucleotides. Ang mga kadena ng nucleotides ay konektado sa pamamagitan ng phosphoric acid residue (phosphodiester bond). Dahil mayroon lamang dalawang uri ng heterocyclic molecule sa nucleotides, ribose at deoxyribose, mayroon lamang dalawang uri ng nucleic acid - deoxyribonucleic acid (DNA) at ribonucleic acid (RNA).
Ang mga monomeric na anyo ay matatagpuan din sa mga cell at may mahalagang papel sa pagbibigay ng senyas o pag-iimbak ng enerhiya. Ang pinakasikat na RNA monomer ay ATP, adenosine triphosphoric acid, ang pinakamahalagang energy accumulator sa cell.
Ang isa sa mga pangunahing tungkulin ng mga nucleic acid ay upang matukoy ang synthesis ng protina. Ang impormasyon tungkol sa istraktura ng mga protina na naka-encode sa pagkakasunud-sunod ng nucleotide ng DNA ay dapat mailipat mula sa isang henerasyon patungo sa isa pa, at samakatuwid ay dapat itong makopya nang tama, i.e. synthesis ng eksaktong parehong molekula ng DNA (pagtitiklop).
№75RepDNA lication ang proseso ng synthesis ng isang molekula ng anak na babae ng deoxyribonucleic acid sa matrix ng molekula ng magulang na DNA. Sa kasunod na paghahati ng selula ng ina, ang bawat cell ng anak na babae ay tumatanggap ng isang kopya ng molekula ng DNA, na kapareho ng DNA ng orihinal na selula ng ina. Tinitiyak ng prosesong ito ang tumpak na paghahatid ng genetic na impormasyon mula sa henerasyon hanggang sa henerasyon. Ang pagtitiklop ng DNA ay isinasagawa ng isang komplikadong enzymatic complex na binubuo ng 15-20 iba't ibang protina, na tinatawag na repisome. Ang pagtitiklop ng DNA ay isang mahalagang kaganapan sa paghahati ng cell. Mahalaga na sa oras ng paghahati ang DNA ay ganap na ginagaya at isang beses lamang. Ito ay ibinibigay ng ilang mga mekanismo ng regulasyon ng pagtitiklop ng DNA. Nagaganap ang pagtitiklop sa tatlong yugto:
pagsisimula ng pagtitiklop
pagpapahaba
pagwawakas ng pagtitiklop.
Ang regulasyon ng pagtitiklop ay isinasagawa pangunahin sa yugto ng pagsisimula. Ito ay medyo madaling ipatupad, dahil ang pagtitiklop ay maaaring magsimula hindi mula sa anumang piraso ng DNA, ngunit mula sa isang mahigpit na tinukoy, na tinatawag na site ng pagsisimula ng pagtitiklop. Maaari lamang magkaroon ng isa o maraming ganoong mga site sa genome. Ang konsepto ng replicon ay malapit na nauugnay sa konsepto ng isang site ng pagsisimula ng pagtitiklop. Ang replicon ay isang kahabaan ng DNA na naglalaman ng site ng pagsisimula ng pagtitiklop at umuulit kapag nagsimula ang DNA synthesis mula sa site na iyon. Ang mga genome ng bakterya, bilang panuntunan, ay kumakatawan sa isang replicon, na nangangahulugang ang pagtitiklop ng buong genome ay resulta ng isang pagkilos lamang ng pagsisimula ng pagtitiklop. Ang mga genome ng mga eukaryote (pati na rin ang kanilang mga indibidwal na chromosome) ay binubuo ng isang malaking bilang ng mga independiyenteng replicon, na makabuluhang binabawasan ang kabuuang oras ng pagtitiklop ng isang indibidwal na kromosoma. Ang mga mekanismo ng molekular na kumokontrol sa bilang ng pagsisimula ng pagtitiklop ay kumikilos sa bawat site sa isang cycle ng cell division ay tinatawag na copy number control. Bilang karagdagan sa chromosomal DNA, ang mga bacterial cell ay kadalasang naglalaman ng mga plasmid, na mga indibidwal na replika. Ang mga plasmid ay may sariling mga mekanismo ng pagkontrol ng kopya: maaari silang magbigay ng synthesis ng isang kopya lamang ng plasmid bawat cell cycle, o libu-libong kopya.
Nagsisimula ang pagtitiklop sa lugar ng pagsisimula ng pagtitiklop na may pag-unwinding ng DNA double helix, na may pagbuo ng isang replication fork - ang site ng direktang pagtitiklop ng DNA. Ang bawat site ay maaaring bumuo ng isa o dalawang replication fork, depende sa kung ang replication ay unidirectional o bi-directional. Mas karaniwan ang bidirectional replication. Ilang oras pagkatapos ng pagsisimula ng pagtitiklop sa isang electron microscope, posibleng obserbahan ang replication eye - isang seksyon ng chromosome kung saan ang DNA ay na-replicate na, na napapalibutan ng mas pinalawig na mga seksyon ng hindi na-replicated na DNA.
Sa replication fork, kinokopya ng DNA ang isang malaking kumplikadong protina (replicisome), ang pangunahing enzyme kung saan ay DNA polymerase. Ang replication fork ay gumagalaw sa bilis na humigit-kumulang 100,000 base pairs kada minuto sa prokaryotes at 500-5,000 sa eukaryotes.
№76 Ang istraktura at mga uri ng chromosome.Ang nucleus ng bawat somatic cell ng katawan ng tao ay naglalaman ng46 chromosome... Kitmga chromosome bawat indibidwal, parehong normal at pathological, ay tinatawag na karyotype.
Mula sa46 chromosomena bumubuo sa human chromosome set, 44 o 22 na pares ang kumakatawan sa autosomalmga chromosome , ang huling pares ay mga sex chromosome. Sa mga kababaihan, ang konstitusyon ng mga sex chromosome ay karaniwang kinakatawan ng dalawang X chromosome, at sa mga lalaki - ng X at Y chromosomes.
Sa lahatmga pares ng chromosome parehong autosomal at kasarian, ang isa sa mga chromosome ay nakuha mula sa ama, at ang isa ay mula sa ina. Ang mga chromosome ng isang pares ay tinatawag na homologues, o homologous chromosome. Ang mga reproductive cell (sperm at itlog) ay naglalaman ng isang haploid set ng mga chromosome, i.e. 23 chromosome.
 Ang tamud ay nahahati sa dalawang uri depende sa kung naglalaman ang mga itokromosomaX o Y. Karaniwang naglalaman ang lahat ng itlogchromosome lamang
X.
Ang tamud ay nahahati sa dalawang uri depende sa kung naglalaman ang mga itokromosomaX o Y. Karaniwang naglalaman ang lahat ng itlogchromosome lamang
X.
Mga Chromosome ay malinaw na nakikita pagkatapos ng espesyal na paglamlam sa panahon ng cell division, kapag ang mga chromosome ay maximally spiralized. Sa kasong ito, may nakitang constriction sa bawat chromosome, na tinatawag na centromere. Hinahati ng centromere ang chromosome sa isang maikling braso (na tinutukoy ng titik "p") at isang mahabang braso (na tinutukoy ng titik na "q"). Tinutukoy ng centromere ang paggalaw ng chromosome sa panahon ng cell division. Ayon sa posisyon ng sentromere, ang mga chromosome ay inuri sa ilang mga grupo. Kung ang centromere ay matatagpuan sa gitna ng chromosome, kung gayon ang naturang chromosome ay tinatawag na metacentric, kung ang centromere ay matatagpuan mas malapit sa isa sa mga dulo ng chromosome, kung gayon ito ay tinatawag na acrocentric. Ang ilang mga acrocentric chromosome ay may tinatawag na mga satellite, na bumubuo ng nucleoli sa isang hindi naghating na selula. Ang nucleoli ay naglalaman ng maraming kopya ng rRNA. Bilang karagdagan, ang mga submetacentric chromosome ay nakikilala, kapag ang centromere ay matatagpuan hindi sa gitna ng chromosome, ngunit medyo lumipat sa isa sa mga dulo, ngunit hindi kasing-kahulugan tulad ng sa acrocentric chromosome.
Ang mga dulo ng bawat isakromosomang balikat tinatawag na telomeres. Napag-alaman na ang mga telomere ay may mahalagang papel sa pagpapanatili ng katatagan ng mga chromosome. Ang mga Telomeres ay naglalaman ng malaking bilang ng mga pag-uulit ng TTAGGG nucleotide sequence, ang tinatawag na tandem repeats. Karaniwan, sa panahon ng paghahati ng cell, mayroong pagbaba sa bilang ng mga pag-uulit na ito mga telomere .
Gayunpaman, sa bawat oras na nakumpleto ang mga ito sa tulong ng isang espesyal na enzyme na tinatawagtelomerase... Ang pagbaba sa aktibidad ng enzyme na ito ay humahantong sa pag-ikli ng telomeres, na pinaniniwalaan na sanhi ng pagkamatay ng cell, at karaniwang sinasamahan ng pagtanda.
№77. Pag-uuri ng chromosome ng Denver. Ang pag-uuri at katawagan ng pare-parehong nabahiran na mga kromosom ng tao ay unang pinagtibay sa isang internasyonal na pagpupulong noong 1960 sa Denver, kalaunan ay medyo binago at dinagdagan (London, 1963 at Chicago, 1966). Ayon sa klasipikasyon ng Denver, ang lahat ng chromosome ng tao ay nahahati sa 7 grupo, na nakaayos sa pagbaba ng pagkakasunud-sunod ng kanilang haba at isinasaalang-alang ang centriole index (ang ratio ng haba ng maikling braso sa haba ng buong chromosome, na ipinahayag bilang isang porsyento). Ang mga pangkat ay itinalaga sa pamamagitan ng mga titik ng alpabetong Ingles mula A hanggang G. Ang lahat ng mga pares ng chromosome ay karaniwang binibilang na may mga Arabic numeral. Ang mga katangian ng mga pangkat ay ipinakita sa talahanayan. 4. Ang iminungkahing pag-uuri ay naging posible upang malinaw na makilala ang mga chromosome na kabilang sa iba't ibang grupo. Mula noong 1960, nagsimula ang mabilis na pag-unlad ng clinical cytogenetics: noong 1959, natuklasan ni J. Lejeune ang chromosomal na katangian ng Down's syndrome; Inilarawan ni K. Ford, P. Jacobs at J. Strong ang mga tampok ng karyotype sa mga sindrom ng Klinefelter at Turner; noong unang bahagi ng 70s. ang chromosomal na katangian ng Edwards at Patau syndromes, ang sindrom ng "sigaw ng pusa" ay natuklasan; inilarawan ang chromosomal instability sa isang bilang ng mga hereditary syndrome at malignant na sakit. Kasabay nito, ang paggamit ng paraan ng pagkuha ng magkakatulad na kulay na mga chromosome ay naging hindi sapat na epektibo para sa pagkilala ng mga chromosome. Ang kawalan ng pag-uuri ng Denver ay ang pagkakaiba-iba ng mga homologous na pares sa loob ng isang pangkat ng mga chromosome ay madalas na nakakaranas ng hindi malulutas na mga paghihirap.
Talahanayan 4^ Mga pangkat ng chromosome at ang kanilang mga katangian
|
Grupo |
Chromosome number |
Lokasyon ng Centromere |
Centriole index (%) |
Tandaan |
|
Ang pinakamalaking metacentric |
Sa mahabang balikat, maaaring mayroong pangalawang paghihigpit |
|||
|
Ang pinakamalaking submetacentric | ||||
|
Malaking metacentric |
20% na mas maikli kaysa sa una |
|||
|
Malaking submetacentric | ||||
|
6-12 at X chromosome |
Katamtamang submetacentric |
Sa ika-9, kadalasang mayroong pangalawang paghihigpit |
||
|
Katamtamang acrocentric |
Sa lahat ng pangalawang paghihigpit |
|||
|
Maliit na metacentric |
Ang pangalawang constriction ay nangyayari sa 10% ng mga kaso. |
|||
|
Maliit na submetacentric | ||||
|
Pinakamaliit na metacentric | ||||
|
21-22 at Y-chromosome |
Pinakamaliit na acrocentric |
Sa ika-21 at ika-22 pangalawang paghihigpit |
№78 Biosynthesis ng protina sa cell. Ang synthesis ng protina ay isang napakakomplikado, maraming hakbang na proseso. Nagaganap ito sa mga espesyal na organelles - ribosomes. Ang cell ay naglalaman ng isang malaking bilang ng mga ribosome. Halimbawa, ang E. coli ay may humigit-kumulang 20,000 sa kanila.
Paano nangyayari ang synthesis ng protina sa mga ribosom?
Ang mga molekula ng protina ay mahalagang mga polypeptide chain na binubuo ng mga indibidwal na amino acid. Ngunit ang mga amino acid ay hindi sapat na aktibo upang magbigkis nang mag-isa. Samakatuwid, bago kumonekta sa isa't isa at bumuo ng isang molekula ng protina, ang mga amino acid ay dapat na maisaaktibo. Ang pag-activate na ito ay nangyayari sa ilalim ng pagkilos ng mga espesyal na enzyme. Bukod dito, ang bawat amino acid ay may sariling enzyme na partikular na nakatutok dito.
Ang pinagmumulan ng enerhiya para dito (tulad ng para sa maraming mga proseso sa cell) ay adenosine triphosphate (ATP).
Bilang resulta ng pag-activate, ang amino acid ay nagiging mas labile at, sa ilalim ng pagkilos ng parehong enzyme, nagbubuklod sa t-RNA.
Mahalaga na ang bawat amino acid ay tumutugma sa isang mahigpit na tiyak na t-RNA. Nahanap niya ang "kanyang" amino acid at inilipat ito sa ribosome. Samakatuwid, ang RNA na ito ay tinawag na transport RNA.
Dahil dito, ang iba't ibang mga aktibong amino acid, kasama ng kanilang mga t-RNA, ay pumapasok sa ribosome. Ang ribosome ay, kumbaga, isang conveyor belt para sa pag-assemble ng isang chain ng protina mula sa iba't ibang mga amino acid na pumapasok dito.
Lumalabas na kasabay ng t-RNA, kung saan "nakaupo" ang sarili nitong amino acid, ang ribosome ay tumatanggap ng "signal" mula sa DNA na nakapaloob sa nucleus. Alinsunod sa signal na ito, ang isa o isa pang protina, ang isa o isa pang enzyme ay synthesize sa ribosome (dahil ang mga enzyme ay mga protina).
Ang direktang impluwensya ng DNA sa synthesis ng protina ay hindi direktang isinasagawa, ngunit sa tulong ng isang espesyal na tagapamagitan, ang anyo ng RNA, na tinatawag na messenger o messenger RNA (m-RNA o i-RNA).
Ang Messenger RNA ay na-synthesize sa nucleus sa pamamagitan ng yodo sa pamamagitan ng impluwensya ng DNA, samakatuwid ang komposisyon nito ay sumasalamin sa komposisyon ng DNA. Ang molekula ng RNA ay parang amag ng anyo ng DNA.
Ang na-synthesize na m-RNA ay pumapasok sa ribosome at, kumbaga, ay naghahatid sa istrukturang ito ng isang plano - sa anong pagkakasunud-sunod ang mga activated amino acid na pumapasok sa ribosome ay dapat pagsamahin sa isa't isa upang ang isang tiyak na protina ay ma-synthesize. Kung hindi, ang genetic na impormasyon na naka-encode sa DNA ay inililipat sa m-RNA at pagkatapos ay sa protina.
№79 Mutation (lat.mutatio - pagbabago) - paulit-ulit (iyon ay, isa na maaaring minana ng mga inapo ng isang naibigay na cell o organismo) na pagbabago sa genotype na nangyayari sa ilalim ng impluwensya ng panlabas o panloob na kapaligiran. Term na likha ni Hugo de Vries. Ang proseso ng paglitaw ng mga mutasyon ay tinatawag na mutagenesis.
Genomic: - polyploidization (ang pagbuo ng mga organismo o mga cell, ang genome nito ay kinakatawan ng higit sa dalawa (3n, 4n, 6n, atbp.) na set ng chromosomes) at neuploidy (heteroploidy) - isang pagbabago sa bilang ng mga chromosome na ay hindi isang multiple ng haploid set (tingnan ang Inge-Vechtomov, 1989). Depende sa pinagmulan ng mga chromosome set, ang polyploid ay nakikilala sa pagitan ng mga allopolyploid, na may mga set ng chromosome na nakuha sa pamamagitan ng hybridization mula sa iba't ibang species, at autopolyploids, na may pagtaas sa bilang ng mga set ng chromosome ng kanilang sariling genome, isang multiple ng n.
Sa mga mutasyon ng chromosomal, nangyayari ang malalaking pagbabago sa istruktura ng mga indibidwal na chromosome. Sa kasong ito, mayroong pagkawala (pagtanggal) o pagdodoble ng isang bahagi (pagdoble) ng genetic na materyal ng isa o ilang mga chromosome, isang pagbabago sa oryentasyon ng mga segment ng chromosome sa mga indibidwal na chromosome (inversion), pati na rin ang paglipat ng isang bahagi ng genetic material mula sa isang chromosome patungo sa isa pa (translocation) (isang matinding kaso - ang unyon ng buong chromosome, ang tinatawag na Robertsonian translocation, na isang transitional option mula sa chromosomal hanggang genomic mutation).
Sa antas ng gene, ang mga pagbabago sa pangunahing istruktura ng DNA ng mga gene sa ilalim ng impluwensya ng mga mutasyon ay hindi gaanong makabuluhan kaysa sa mga chromosomal mutations, gayunpaman, ang mga mutation ng gene ay mas karaniwan. Bilang resulta ng mga mutation ng gene, mga pagpapalit, pagtanggal at pagpasok ng isa o higit pang mga nucleotide, nagaganap ang mga pagsasalin, pagdoble at pagbabaligtad ng iba't ibang bahagi ng gene. Sa kaso kapag isang nucleotide lamang ang nagbabago sa ilalim ng pagkilos ng isang mutation, ang isa ay nagsasalita ng point mutations. Dahil ang DNA ay naglalaman lamang ng dalawang uri ng nitrogenous base - purines at pyrimidines, ang lahat ng point mutations na may base substitutions ay nahahati sa dalawang klase: transition (pagpapalit ng purine ng purine o pyrimidine na may pyrimidine) at transversion (pagpapalit ng purine ng pyrimidine o vice versa) . Mayroong apat na posibleng genetic na kahihinatnan ng point mutations: 1) pagpapanatili ng kahulugan ng codon dahil sa pagkabulok ng genetic code (synonymous termination (nonsense mutation). Mayroong tatlong walang kahulugan na codon sa genetic code: amber - UAG, ocp - UAA at opal - UGA (alinsunod dito, ang pangalan at mutasyon na humahantong sa pagbuo ng mga walang kahulugan na triplets - halimbawa, amber mutation), 4) reverse pagpapalit (stop codon na may sense codon).
Ayon sa kanilang impluwensya sa pagpapahayag ng gene, ang mga mutasyon ay nahahati sa dalawang kategorya: mga mutasyon ng uri ng mga pagpapalit ng pares ng base at ang uri ng paglilipat ng frame (frameshift). Ang huli ay ang mga pagtanggal o pagpapasok ng mga nucleotide, na ang bilang nito ay hindi isang multiple ng tatlo, na nauugnay sa triplet na katangian ng genetic code.
Ang pangunahing mutation ay tinatawag minsan na direktang mutation, at ang mutation na nagpapanumbalik sa orihinal na istruktura ng isang gene ay tinatawag na reverse mutation, o reversion. Ang pagbabalik sa orihinal na phenotype ng mutant organism dahil sa pagpapanumbalik ng function ng mutant gene ay kadalasang nangyayari hindi dahil sa isang tunay na reversion, ngunit dahil sa isang mutation sa ibang bahagi ng parehong gene o kahit na isa pang non-allelic gene. Sa kasong ito, ang paulit-ulit na mutation ay tinatawag na suppressor mutation. Ang mga genetic na mekanismo kung saan ang mutant phenotype ay pinigilan ay napaka-magkakaibang.
Ang mga mutation sa bato (spores) ay mga persistent somatic mutations na nagaganap sa mga cell ng mga growth point ng halaman. Humantong sa clonal variation. Ang mga ito ay pinapanatili sa panahon ng vegetative propagation. Maraming mga cultivars ay kidney mutations.
№80. Paglalagay ng DNA sa isang chromosome. Ang mga chromosome ay binubuo ng chromatin - isang pinaghalong DNA at mga protina sa isang ~ 1: 1 ratio. Ito ay dahil sa pakikipag-ugnayan sa mga protina na ang molekula ng DNA ay may isang compact na istraktura. Ang average na haba ng isang strand ng DNA ng tao (kung nakaunat) ay 4 cm. Ibig sabihin, ang nucleus ng bawat cell ay naglalaman ng humigit-kumulang 1 metro ng DNA. Ang pag-iimpake ng molekula ng DNA sa chromosome ay may tatlong antas (tingnan ang figure): 1.nucleosome, nucleosome strand 2. solenoid helix 3. mga loop Ang double DNA helix na kilala sa bawat isa sa mga uri nito ay "nakapulupot" sa paligid ng isang complex ng 4 na pares ng ang mga molekula ng protina ay bumubuo ng isang "bead" - nucleosome. Ang mga nucleosome, na magkakaugnay ng mga seksyon ng molekula ng DNA, ay bumubuo ng isang thread, na kung saan ay baluktot sa anyo ng isang solenoid (ito ay ayon sa isa sa mga umiiral na mga modelo), isang pagliko - 6 na mga nucleosome. Ang istraktura na ito ay bumubuo ng mga loop sa susunod na antas ng packaging. Sa form na ito, ang DNA ay naroroon sa cell nucleus sa yugto ng paglaki ng cell, synthesis ng mga bahagi nito, kabilang ang pagdoble ng DNA. Gayunpaman, sa iba't ibang yugto ng buhay ng cell, iba ang chromatin condensation. Sa panahon ng paglaki ng cell, ang chromatin sa nucleus ay hindi na-condensed, ang compactness ng lokasyon nito ay naiiba para sa iba't ibang bahagi ng chromosome. Kapag nangyari ang proseso ng paghahati ng cell, ang chromatin ay nagiging mas siksik at ang mga loop, na nabanggit sa itaas, ay bumubuo ng "mga rosette" na mahigpit na nakaimpake sa isa't isa (ayon din sa isa sa mga umiiral na mga modelo), 18 na mga loop bawat singsing. Bilang resulta, ang chromatin ay nasa anyo ng mga condensed chromosome, na kilala sa atin mula sa mga aklat-aralin.
№81. Gametogenesis.(mula sa Gamete at Greek genesis - pinanggalingan) ang proseso ng pag-unlad at pagbuo ng mga sex cell - gametes (Tingnan ang Gametes). G. male gametes (spermatozoa, sperm) ay tinatawag na spermatogenesis, female gametes (oocytes) - oogenesis. Sa mga hayop at halaman, ang G. ay nagpapatuloy nang iba, depende sa lugar ng meiosis sa siklo ng buhay ng mga organismong ito.
Sa mga multicellular na hayop, ang G. ay nangyayari sa mga espesyal na organo - ang gonads, o gonads (ovaries, testes, at hermaphroditic gonads), at binubuo ng tatlong pangunahing yugto: 1) pagpaparami ng mga pangunahing germ cell - gametogonia (spermatogonia at oogonia) sa pamamagitan ng isang serye ng sunud-sunod na mitoses. 2) ang paglaki at pagkahinog ng mga selulang ito ay tinatawag na ngayong mga gametocytes (spermatocytes at oocytes), na, tulad ng gametogonia, ay mayroong kumpletong (karamihan ay diploid) na hanay ng mga chromosome (tingnan ang Chromosome). Sa oras na ito, ang pangunahing kaganapan ng G. sa mga hayop ay nangyayari - ang dibisyon ng mga gametocytes sa pamamagitan ng meiosis, na humahantong sa isang pagbawas (halving) ng bilang ng mga chromosome sa mga cell na ito at ang kanilang pagbabago sa mga haploid cell (tingnan ang Haploid) - spermatids at ootids ; 3) ang pagbuo ng tamud (o tamud) at mga itlog; sa kasong ito, ang mga itlog ay binibihisan ng isang bilang ng mga embryonic membrane, at ang spermatozoa ay nakakakuha ng flagella, na tinitiyak ang kanilang kadaliang kumilos. Sa mga babae ng maraming species ng hayop, ang meiosis at pagbuo ng itlog ay nakumpleto pagkatapos ng pagtagos ng tamud sa cytoplasm ng oocyte, ngunit bago ang pagsasanib ng nuclei ng tamud at ng itlog.
Sa mga halaman, ang G. ay nahihiwalay sa meiosis at nagsisimula sa mga haploid na selula, sa mga spores (sa mas matataas na halaman, microspores at megaspores). Mula sa mga spores, ang sekswal na henerasyon ng halaman ay bubuo - ang haploid gametophyte, sa mga maselang bahagi ng katawan kung saan - gametangia (lalaki - antheridia (tingnan ang Anteridium), babae - archegonia (Tingnan ang Archegonia)) sa pamamagitan ng mitosis, G. Ang pagbubukod ay gymnosperms at angiosperms, kung saan ang spermatogenesis ay direktang napupunta sa germinating microspore - ang pollen cell. Sa lahat ng mas mababa at mas mataas na spore halaman G. sa antheridia ay isang maramihang dibisyon ng mga cell, bilang isang resulta kung saan ang isang malaking bilang ng mga maliliit na motile spermatozoa ay nabuo. G. sa archegonia - ang pagbuo ng isa, dalawa o higit pang mga oocytes. Sa gymnosperms at angiosperms, ang male G. ay binubuo ng paghahati (sa pamamagitan ng mitosis) ang nucleus ng pollen cell sa generative at vegetative at higit pang paghahati (din sa pamamagitan ng mitosis) ng generative nucleus sa dalawang sperm. Ang paghahati na ito ay nagaganap sa tumutubo na pollen tube. Ang Female G. sa angiosperms ay ang paghihiwalay sa pamamagitan ng mitosis ng isang egg cell sa loob ng isang walong-nucleus embryo sac. Ang pangunahing pagkakaiba ni G. sa mga hayop at halaman: sa mga hayop, pinagsasama nito ang pagbabago ng mga selula mula sa diploid patungo sa haploid at ang pagbuo ng mga haploid gametes; sa mga halaman, ang G. ay nabawasan sa pagbuo ng mga gametes mula sa mga haploid cells.
№82 Karyotyp - isang hanay ng mga tampok (bilang, laki, hugis, atbp.) ng isang kumpletong hanay ng mga chromosome na likas sa mga cell ng isang partikular na biological species (species karyotype), isang partikular na organismo (indibidwal na karyotype) o linya (clone) ng mga cell. Ang visual na representasyon ng isang kumpletong hanay ng chromosome (karyogram) ay tinatawag ding karyotype. Ang hitsura ng mga chromosome ay makabuluhang nagbabago sa panahon ng cell cycle: sa panahon ng interphase, ang mga chromosome ay naisalokal sa nucleus, bilang isang panuntunan, despiralized at mahirap na obserbahan, samakatuwid, ang mga cell ay ginagamit upang matukoy ang karyotype sa isa sa mga yugto ng kanilang dibisyon - mitosis metaphase. Para sa pamamaraan para sa pagtukoy ng karyotype, maaaring gamitin ang anumang populasyon ng naghahati na mga selula. Upang matukoy ang karyotype ng tao, bilang panuntunan, ginagamit ang mga peripheral blood lymphocytes, ang paglipat kung saan mula sa resting stage G0 hanggang sa paglaganap ay pinukaw ng pagdaragdag ng mitogenaphytohemagglutinin. Ang mga selula ng utak ng buto o pangunahing kultura ng mga fibroblast ng balat ay maaari ding gamitin upang matukoy ang karyotype. Upang madagdagan ang bilang ng mga cell sa yugto ng metaphase, ang colchicinilinocadazole ay idinagdag sa kultura ng cell sa ilang sandali bago ang pag-aayos, na humahadlang sa pagbuo ng mga microtubule, at sa gayon ay pinipigilan ang mga chromatids na mag-diver sa mga pole ng cell division at ang pagkumpleto ng mitosis.
Pagkatapos ng pag-aayos, ang mga paghahanda ng metaphase chromosome ay nabahiran at nakuhanan ng litrato; ang tinatawag na sistematikong karyotype ay nabuo mula sa mga micrograph - isang may bilang na hanay ng mga pares ng homologous chromosome, ang mga imahe ng chromosome ay naka-orient nang patayo na may maiikling mga braso pataas, ang kanilang pagnunumero ay ginagawa sa pagpapababa ng pagkakasunud-sunod ng laki, isang pares ng sex chromosome ay inilalagay sa ang dulo ng set (tingnan ang Fig. 1).
Sa kasaysayan, ang unang hindi detalyadong mga karyotypes na naging posible upang pag-uri-uriin ayon sa morpolohiya ng mga chromosome ay nakuha sa pamamagitan ng paglamlam ng Romanovsky-Giemsa, gayunpaman, ang karagdagang pagdedetalye ng istraktura ng mga chromosome sa mga karyotype ay naging posible sa pagdating ng mga diskarte sa paglamlam ng differential chromosome. Ang pinakakaraniwang ginagamit na pamamaraan sa medikal na genetika ay ang G-differential chromosome staining.
Ang paghahambing ng mga complex ng transverse mark sa classical na karyotypy o mga rehiyon na may mga tiyak na spectral na katangian ay ginagawang posible upang makilala ang parehong homologous chromosomes at ang kanilang mga indibidwal na rehiyon, na ginagawang posible upang matukoy nang detalyado ang mga chromosomal aberrations - intra- at interchromosomal rearrangements, na sinamahan ng isang paglabag ng pagkakasunud-sunod ng mga fragment ng chromosome (pagtanggal, pagdoble, pagbabaligtad, pagsasalin). Ang ganitong pagsusuri ay may malaking kahalagahan sa medikal na kasanayan, na nagpapahintulot sa isa na masuri ang isang bilang ng mga chromosomal na sakit na sanhi ng parehong matinding paglabag sa mga karyotypes (mga abnormalidad sa bilang ng mga chromosome) at isang paglabag sa istruktura ng chromosomal o multiplicity ng mga cellular karyotypes sa katawan (mosaicism).
№83 Meiosis- reduction division, maturation division, isang paraan ng cell division, bilang isang resulta kung saan mayroong pagbaba (pagbawas) sa bilang ng mga chromosome ng kalahati at isang diploid cell (naglalaman ng dalawang set ng chromosome) pagkatapos ng dalawang mabilis na sumusunod sa isa't isa Ang mga dibisyon ay nagbubunga ng 4 na haploid (naglalaman ng isang hanay ng mga chromosome).
Biyolohikal na halaga M ... ay upang mapanatili ang pagkakapare-parehoKaryotype at sa isang bilang ng mga henerasyon ng mga organismo ng ganitong uri at tinitiyak ang posibilidad ng recombination) ng mga chromosome at gene sa panahon ng sekswal na proseso. M. ay isa sa mga pangunahing mekanismo ng pagmamana
Ang Meiosis ay binubuo ng 2 magkakasunod na dibisyon na may maikling interphase sa pagitan ng mga ito.
Prophase I - ang prophase ng unang dibisyon ay napaka kumplikado at binubuo ng 5 yugto:
Leptotene o leptoneme - pag-iimpake ng mga kromosom, paghalay ng DNA na may pagbuo ng mga kromosom sa anyo ng mga manipis na mga thread (pinaikli ang mga kromosom).
Zygotene o zigoneme - nangyayari ang conjugation - ang kumbinasyon ng mga homologous chromosome na may pagbuo ng mga istruktura na binubuo ng dalawang konektadong chromosome, na tinatawag na tetrads o bivalents, at ang kanilang karagdagang compactization.
Pachytene o pachinema - (ang pinakamahabang yugto) - sa ilang mga lugar, ang mga homologous chromosome ay mahigpit na konektado, na bumubuo ng chiasmata. Sa kanila meron tumatawid- pagpapalitan ng mga site sa pagitan ng mga homologous chromosome.
Diplotene o Diploneme - ang bahagyang decondensation ng mga chromosome ay nangyayari, habang ang bahagi ng genome ay maaaring gumana, ang mga proseso ng transkripsyon (RNA formation), pagsasalin (protein synthesis) ay nagaganap; Ang mga homologous chromosome ay nananatiling konektado sa isa't isa. Sa ilang mga hayop, ang mga chromosome sa mga oocytes sa yugtong ito ng meiotic prophase ay nakakakuha ng isang katangian na anyo lamp brush chromosomes.
Diakinesis - Nag-condensed muli ang DNA hangga't maaari, huminto ang mga sintetikong proseso, natutunaw ang nuclear shell; centrioles diverge sa pole; Ang mga homologous chromosome ay nananatiling konektado sa isa't isa.
Sa pagtatapos ng Prophase I, ang mga centriole ay lumilipat sa mga pole ng cell, ang fission spindle filament ay nabuo, ang nuclear membrane at nucleoli ay nawasak.
Metaphase I - ang mga bivalent chromosome ay nakahanay sa kahabaan ng ekwador ng cell.
Anaphase I - ang microtubule ay nagkontrata, ang mga bivalents ay naghahati at ang mga kromosom ay naghihiwalay sa mga pole. Mahalagang tandaan na, dahil sa conjugation ng mga chromosome sa zygotene, ang buong chromosome ay naghihiwalay sa mga pole, bawat isa ay binubuo ng dalawang chromatids, at hindi hiwalay na chromatids, tulad ng sa mitosis.
Telophase I - ang mga chromosome ay despiralized at isang nuclear envelope ang lilitaw.
Ang pangalawang dibisyon ng meiosis ay agad na sumusunod sa una, nang walang binibigkas na interphase: ang S-period ay wala, dahil walang pagtitiklop ng DNA bago ang pangalawang dibisyon.
Prophase II - ang condensation ng chromosomes ay nangyayari, ang cell center ay nahahati at ang mga produkto ng dibisyon nito ay naghihiwalay sa mga pole ng nucleus, ang nuclear envelope ay nawasak, at isang fission spindle ay nabuo.
Metaphase II - univalent chromosomes (bawat isa ay binubuo ng dalawang chromatids) ay matatagpuan sa "equator" (sa pantay na distansya mula sa "poles" ng nucleus) sa isang eroplano, na bumubuo ng tinatawag na metaphase plate.
Anaphase II - univalents divide atchromatidlumihis sa mga poste.
Telophase II - ang mga chromosome ay despiralized at isang nuclear envelope ang lilitaw.
Bilang resulta, mula sa isadiploid na selulaapatmga haploid na selula... Sa mga kaso kung saan ang meiosis ay nauugnay sagametogenesis(halimbawa, sa mga multicellular na hayop), sa panahon ng pag-unladmga oocytes ang una at pangalawang dibisyon ng meiosis ay hindi pantay. Bilang isang resulta, isang haploid na itlog at tatlong tinatawag na mga katawan ng pagbabawas(mga abortive derivatives ng una at pangalawang dibisyon).
№84 Dibisyon ng cell - mitosis
Kung naobserbahan mo ang cell division sa ilalim ng mikroskopyo, makikita mo ang mga sumusunod. Una, ang materyal sa loob ng core ay nagiging napaka siksik. Maaari mo ring makilala sa pagitan ng mga discrete na istruktura - mga chromosome. Alam mo na ang mga selulang somatic (non-sex) ng tao ay may 23 pares ng chromosome - isang chromosome mula sa bawat pares mula sa bawat magulang. Sa kabuuan, mayroong 46 chromosome sa mga somatic cell ng tao. Lahat ng 46 ay pumila sa equatorial plane ng cell. Sa yugtong ito, malinaw na nakikita na ang bawat isa sa 46 na chromosome ay aktwal na binubuo ng dalawang chromatids, na konektado sa isa't isa sa isang rehiyon na katulad ng isang pindutan. Ang lugar na ito ay tinatawag na sentromere. Sa magkabilang dulo ng cell, mayroong dalawang siksik na mga spot (centrioles), na ang bawat isa ay bumubuo ng mga thread ng division spindle. Ang bawat strand ay sumasali sa chromosome sa sentromere. Dalawang strand ang nakakabit sa bawat pares ng chromosome - isa mula sa bawat centriole. Pagkatapos ay makikita natin na ang mga centrioles, tulad ng mga coils, ay nagpapaikut-ikot sa mga thread ng fission spindle, bilang isang resulta kung saan ang mga chromosome ay diverge sa mga pole ng cell. Pagkatapos ay hinati ang mga nilalaman ng cell. Ito ay isang kamangha-manghang tanawin.
Ang mga kaganapan ay nabuo ayon sa sumusunod na senaryo. Ito ay kinakailangan upang maunawaan na para sa cell division isang napakahalagang gawain ay dapat malutas - DNA pagdoble (pagtitiklop) ay dapat mangyari, dahil ito ay sa DNA na ang lahat ng impormasyon tungkol sa kung ano at paano dapat mangyari sa cell ay naka-encode. Tandaan na ang DNA ay isang double helix. Sa labas ng spiral ay ang sugar-phosphate backbone, kung saan ang mga sugars ay naka-link sa isa't isa sa pamamagitan ng covalent bonds sa pamamagitan ng phosphoric acid residue. Sa loob ng spiral mayroong mga organikong nitrogenous na base na konektado sa mga pares ng mga bono ng hydrogen. Ang kahulugan ng pagkopya ng impormasyon na nakaimbak sa isang chromosome at paglilipat ng impormasyong ito sa isang bagong cell nang walang mga pagbabago ay napakasimple: kinakailangang i-unwind ang double helix ng DNA at putulin ang mga hydrogen bond na humahawak sa mga pares ng nitrogenous base na magkasama. Mula sa sandaling ito, lumilitaw ang dalawang solong chain na may nitrogenous base, na nakausli mula sa backbone ng asukal-pospeyt. Tandaan na ang bawat nitrogenous base ng isa sa mga DNA strand ay palaging bumubuo ng mga bond na may napakaspesipikong base ng isa pang DNA strand. Kaya, ang adenine ay nagbubuklod lamang sa thymine, at ang guanine lamang sa cytosine. Ito ang prinsipyo ng complementarity ng nitrogenous bases. Tandaan din na ang mga nucleotide ay mga molekula na binubuo ng isang organic na base at phosphorylated na asukal. Sa teorya, kung ang mga kinakailangang nucleotide ay naroroon sa nucleoplasm, ang bawat nitrogen base ng bawat DNA strand ay tutugma sa sarili nito sa isang angkop na pares. Mayroong tiyak at napakahalagang enzyme, ang DNA polymerase, na ginagawang posible ang reaksyong ito. Ang mga asukal ng bagong nabuo na DNA strand ng anak na babae ay konektado sa isa't isa sa pamamagitan ng mga phosphodiester bond, bilang isang resulta kung saan nabuo ang isang sugar-phosphate backbone - at nangyari ito! Mayroon kang dalawang magkaparehong chromosome kung saan nagkaroon ka lang ng isa (Figure 2.7). Siyempre, sa katunayan, ang proseso ng pagtitiklop ng molekula ng DNA sa isang cell ay mas kumplikado at depende sa paggana ng isang bilang ng mga enzyme, ngunit pag-uusapan natin ito sa ibaba. Ngayon tingnan natin ang proseso ng paghahati ng selula. Sa una, ang mga pares ng chromosome ay nasa despiralized na estado sa nucleus at hindi nakikilala sa ilalim ng mikroskopyo. Ang nasabing mga cell ay sinasabing nasa interphase. Sa mga ito, ang genetic na materyal na kilala bilang chromatin ay isang gusot na masa at hindi kasing siksik tulad ng kapag ang mga chromosome ay nakikilala sa ilalim ng mikroskopyo. Nakuha ng Chromatin ang pangalan nito mula sa katotohanan na ito ay nabahiran ng madilim na may mga espesyal na kulay na tina. Nangyari ito nang matagal bago malaman ng mga tao ang tungkol sa tunay na paggana nito. Tulad ng nabanggit na, ang mga somatic cell ng tao ay may 23 pares ng chromosome. Dalawampu't dalawang pares ang tinatawag na autosomal chromosomes, na pareho sa mga lalaki at babae. Ang dalawampu't tatlong pares ay kinakatawan ng mga chromosome ng sex, na iba sa mga lalaki at babae. Ang mga babae ay may dalawang malalaking chromosome na tinatawag na X chromosome, habang ang mga lalaki ay may isang malaking X chromosome at isang maliit na Y chromosome. Ang mga chromosome ng isang pares ay may parehong istraktura. Gayunpaman, ang mga chromosome na natanggap mula sa iba't ibang mga magulang ay nagdadala ng iba't ibang impormasyon, at sa ganitong kahulugan, ang mga pares ng chromosome ay hindi magkapareho.
Ang mitosis na ipinapakita sa Fig. 2.8, ay isang unibersal na paraan ng paghahati ng anumang eukaryotic cells. Ang mitosis ay binubuo ng apat na pangunahing natatanging mga yugto: prophase, metaphase, anaphase, telophase. Kapag nagsimulang maghati ang isang cell, nadoble na ang genetic material ng bawat chromosome. Sa prophase, ang bilang ng mga chromosome ay tumutugma sa 46, ngunit ang bawat chromosome ay doble, iyon ay, ito ay binubuo ng dalawang kapatid na chromatids.
Upang simulan ang pagdoble ng genetic na materyal, ang cell ay kailangang lutasin ang isang mahirap na gawain - upang i-unwind ang DNA helix. Karamihan sa atin ay nag-iisip na ang chromosome ay hindi ganoon kahaba, dahil ganoon ang nakasanayan nating makita ang mga ito sa lahat ng mga guhit. Sa katotohanan, sa isang interphase cell, ang molekula ng DNA ay nasa isang supercoiled na estado, iyon ay, ito ay napilipit nang mahigpit at sa isang tiyak na pagkakasunud-sunod. Ang indikasyon ay ang katotohanan na kung ang molekula ng DNA ay nasa loob ng selula sa isang untwisted na estado, ang haba nito ay magiging mga 1 m! Isipin na lang kung gaano kakomplikado at kaayusan ang pag-iimpake ng napakaraming genetic material sa loob ng isang microscopic cell. Mula dito nagiging malinaw kung bakit sa mga eukaryotic cell ang double helix ng molekula ng DNA ay naka-pack sa isang compact na istraktura na nabuo sa pamamagitan ng pakikipag-ugnayan sa isang bilang ng mga protina na tinatawag na histones. Kaya, ang mga chromosomal na protina ay nag-iimpake ng higanteng molekula sa isang nucleus na may dami lamang ng ilang cubic microns (Larawan 2.9) Kaagad bago ang simula ng proseso ng pagtitiklop, ang molekula ng DNA ay natanggal sa sugat ng enzyme helicase. Pagkatapos ay nasira ang mga bono ng hydrogen, naghihiwalay ang mga kadena, at ang mga kadena ng anak na babae ay nabuo sa kanilang matris ayon sa prinsipyo ng complementarity. Ang paghahati ng cell ay mahigpit na kinokontrol. May mga cell na madalas na naghahati. Kabilang dito ang mga selula ng bone marrow. Ang iba pang mga selula, tulad ng mga selula ng buto, ay nahahati pangunahin sa panahon ng aktibong paglaki o pagkasira ng organ. Sa wakas, may mga cell na napakabihirang hatiin o hindi talaga. Kasama sa ganitong uri ang mga mature na nerve cells. Ang hindi makontrol na paghahati at paglaki ng cell ay isang tanda ng kanilang pagkabulok ng kanser. Ang proseso ng cell division ay nangangailangan ng mga regulator na tinatawag na activation factor. Ang mga kadahilanan ng paglago ay mga halimbawa ng mga naturang pabrika. Ang pag-aaral ng mga kadahilanan ng paglago ay isa sa pinakamabilis na lumalagong mga lugar sa intersection ng modernong biology at medisina. Maraming mga kadahilanan ang kasalukuyang naka-highlight at bahagyang nailalarawan.
Karaniwang gumagana ang mga activation factor sa pamamagitan ng pagbabago ng hugis ng DNA at pagpapahintulot sa DNA polymerase, na responsable para sa pagtitiklop ng DNA, na magbigkis dito. Ang DNA polymerase ay unang nakahanap ng isang tiyak na site sa molekula ng DNA. Ang hugis-loop na rehiyon na ito ay kinakailangan para sa DNA polymerase na magbigkis sa DNA. Sa turn, ito ay nabuo kapag ang activation factor ay nagbubuklod sa DNA. Ang lugar ng molekula ng DNA kung saan nakakabit ang DNA polymerase at nagsisimula ang proseso ng pagtitiklop ay tinatawag na punto ng pinagmulan ng pagtitiklop. Dapat mo ring malaman na ang mga maikling RNA sequence ay kinakailangan upang simulan ang DNA synthesis, na gumagana bilang "primer". Ang pagkakaroon ng nakatali sa isang molekula ng DNA, ang DNA polymerase ay gumagalaw sa kahabaan ng maternal chain, na nakumpleto ang pagtatayo ng chain ng anak na babae ayon sa prinsipyo ng complementarity.
Ang isa sa pinakamahalagang tungkulin ng DNA polymerase ay ang pagwawasto ng mga error sa bagong synthesize na DNA strand. Ang pagkilala sa mga maling pares ng base, ang DNA polymerase ay babalik ng isang hakbang, ang maling base ay tinanggal mula sa anak na hibla, at ang tama ay ipinasok. Bilang resulta, ang bilang ng mga error sa pagtitiklop ng DNA ay humigit-kumulang isang error sa 10-100 bilyong base pairs. Gayunpaman, dahil ang mga cell ay madalas na naghahati, ang mga error ay hindi maiiwasang maipon. Marami sa mga error na ito ay nakakaapekto sa cell viability. Kung ang mga selula kung saan naganap ang mutation, gayunpaman ay nabubuhay, sinisira sila ng immune system sa tulong ng mga espesyal na selula na inangkop para sa layuning ito.
Hayaan akong bumalik ngayon sa proseso ng cell division. Sa prophase ng mitosis, ang mga chromosome ay binubuo ng dalawang chromatids, na resulta ng pagtitiklop ng DNA sa interphase. Gayunpaman, hindi posible na irehistro ang mga ito sa ilalim ng isang mikroskopyo, dahil sa prophase ang mga kapatid na chromatids ay malapit na nakikipag-ugnayan sa isa't isa, na magkaparehong umiikot sa isang kamag-anak sa isa. Sa prometaphase, ang nuclear membrane ay nawasak, at ang mga chromosome ay nagsisimulang lumipat patungo sa equatorial plane ng cell. Tinitiyak ng prosesong ito ang normal na paghihiwalay ng mga chromosome sa mga pole ng cell sa panahon ng cell division, bilang isang resulta kung saan ang isang kumpletong hanay ng mga chromosome ay napupunta sa bawat isa sa mga bagong nabuo na mga cell. Ang mga kapatid na chromatids ng isang chromosome ay naka-link sa isa't isa sa isang punto na tinatawag na sentromere. Sa lugar ng centromere, mayroong isang kinetochore, isang kumplikadong istraktura ng protina na nauugnay sa DNA ng sentromeric na rehiyon. Ang mga filament ng fission spindle ay nakakabit sa zone na ito sa panahon ng mitosis. Tulad ng naobserbahan mo na, ang mga centriole ay matatagpuan sa mga pole ng cell, ang bilang nito ay dumoble din bago ang cell division. Ang mga centriole ay kasangkot sa pagbuo ng mga filament ng fission spindle. Ang bawat spindle filament ay nakakabit sa isang chromosome sa rehiyon ng kinetochore. Sa ikalawang yugto ng mitosis, metaphase, chromosome, na binubuo ng dalawang chromatids, ay nakahanay sa equatorial plane ng cell.
Ang mga filament ng fission spindle ay humihila ng mga chromosome, na tinawag mula sa sandaling ito na mga anak na chromosome, sa magkabilang dulo ng cell. Ang yugto kung saan gumagalaw ang mga kromosom sa iba't ibang direksyon ay tinatawag na anaphase. Kung ang lahat ay naaayon sa plano, ang centriole ay naghahati, at ang mga kapatid na chromatids ay naghihiwalay sa mga pole ng cell. Kung ang mga chromosome ay hindi pumila sa equatorial plane ng cell sa wastong pagkakasunud-sunod, ang proseso ng cell division ay maaaring maputol.
Matapos kumalat ang mga chromosome sa magkabilang dulo ng parent cell, nahahati ang cell sa dalawa bilang resulta ng cytokinesis. Ang ikaapat at huling yugto ng mitosis ay tinatawag na telophase. Ang mga cell na dumarami sa pamamagitan ng mitosis, tulad ng nabanggit na, ay tinatawag na somatic. Kabilang dito ang lahat ng mga selula sa ating katawan, maliban sa mga selula na kalaunan ay naging tamud o itlog. Ang mga selula na nag-aambag sa pagbuo ng isang bagong organismo ay tinatawag na mga selulang mikrobyo. Ang mga sex cell ay nabuo bilang isang resulta ng meiosis, na isasaalang-alang natin sa susunod na seksyon.
Ang mitosis ay kadalasang nalulutas nang walang pagkaantala. Gayunpaman, sa anumang naibigay na sandali, milyon-milyong mga cell division ang nagaganap sa katawan ng tao. Paminsan-minsan, mayroong masyadong maraming genetic na materyal sa mga cell, habang sa iba ay maaaring mas mababa ito kaysa sa normal. Karaniwan, ang mga selulang ito ay hindi nabubuhay o inaalis ng immune system.
№85 Mga namamana na sakit. Pag-uuri ng mga namamana na sakit. Mga namamana na sakit - mga sakit, ang paglitaw at pag-unlad nito ay nauugnay sa mga depekto sa software ng mga cell, na minana sa pamamagitan ng mga gametes. Ang termino ay ginagamit na may kaugnayan sa polyetiological na mga sakit, sa kaibahan sa isang mas makitid na grupo - mga genetic na sakit.
Depende sa uri ng pangunahing apektadong mga selula, ang mga sumusunod na grupo ng mga sakit ay nakilala: mga sakit dahil sa mga mutasyon sa mga selula ng mikrobyo - "gametic", i.e. aktwal na namamana na mga sakit (halimbawa, phenylketonuria, hemophilia); ang mga sakit na ito ay namamana; sakit dahil sa mutations sa somatic cells - "somatic" (halimbawa, mga tumor, ilang mga sakit ng immune autoaggression). Ang mga sakit na ito ay hindi namamana; mga sakit dahil sa kumbinasyon ng mga mutasyon sa mikrobyo at somatic cells (halimbawa, familial retinoblastoma).
Ang mga nakamamatay, sublethal at hypogenital na sakit ay isinasaalang-alang nang hiwalay: ang mga nakamamatay na sakit ay humahantong sa kamatayan sa panahon ng intrauterine development (halimbawa, monosomy ng autosomes, haploidy, karamihan sa polyploidy); Ang mga sublethal na sakit ay humahantong sa pagkamatay ng indibidwal bago ang pagdadalaga (halimbawa, hereditary immunodeficiencies tulad ng Swiss-type na agam-maglobulinemia, Louis-Bar syndrome, ilang hemophilia); Ang mga hypogenital na sakit ay pinagsama sa kawalan ng katabaan (halimbawa, Shereshevsky-Turner, Kleinfelter syndromes).
Depende sa papel na ginagampanan ng pagmamana at kapaligiran, apat na mga grupo ng mga sakit ay nakikilala: namamana sakit tamang (bumuo bilang isang resulta ng mutations); mga sakit sa ecogenetic (bumuo sa pagkakaroon ng isang mutation sa ilalim ng impluwensya ng isang tiyak na kadahilanan sa kapaligiran); mga sakit ng genetic predisposition (halata ang kontribusyon ng genetic factor); mga sakit na sanhi ng mga kadahilanan sa kapaligiran (mahigpit na nagsasalita, ang pangkat ng mga sakit na ito ay hindi nabibilang sa mga namamana na sakit, dahil ang kontribusyon ng mga genetic na kadahilanan sa kanilang paglitaw, pag-unlad at kinalabasan ay maaaring napakaliit, nagdududa o hindi alam).